

Insula and Sylvian Fissure

This article was originally published here: Tanriover N, Rhoton AL Jr., Kawashima M, Ulm AJ, Yasuda A: Microsurgical anatomy of the insula and the sylvian fissure. J Neurosurg 100:891–922, 2004 and is included through an exclusive partnership with the Journal of Neurosurgery and its parent company, the American Association of Neurological Surgeons (AANS). The AANS retains full copyright. The appearance of this material here does not imply open access or free use by any other party.

Atlas Choice Tapered Pattie Collection
Low-profile for maximal visualization and protection
Tapered shape designed for retractorless surgery
Unparalleled flexibility and non-stick features
Abstract
OBJECT The purpose of this study was to define the topographic anatomy, arterial supply, and venous drainage of the insula and sylvian fissure.
METHODS The neural, arterial, and venous anatomy of the insula and sylvian fissure were examined in 43 cerebral hemispheres.
CONCLUSIONS The majority of gyri and sulci of the frontoparietal and temporal opercula had a constant relationship to the insular gyri and sulci and provided landmarks for approaching different parts of the insula. The most lateral lenticulostriate artery, an important landmark in insular surgery, arose 14.6 mm from the apex of the insula and penetrated the anterior perforated substance 15.3 mm medial to the limen insulae. The superior trunk of the middle cerebral artery (MCA) and its branches supplied the anterior, middle, and posterior short gyri; the anterior limiting sulcus; the short sulci; and the insular apex. The inferior trunk supplied the posterior long gyrus, inferior limiting sulcus, and limen area in most hemispheres. Both of these trunks frequently contributed to the supply of the central insular sulcus and the anterior long gyrus. The areas of insular supply of the superior and inferior trunks did not overlap. The most constant insular area of supply by the cortical MCA branches was from the prefrontal and precentral arteries that supplied the anterior and middle short gyri, respectively. The largest insular perforating arteries usually arose from the central and angular arteries and most commonly entered the posterior half of the central insular sulcus and posterior long gyrus. Insular veins drained predominantly to the deep middle cerebral vein, although frequent connections to the superficial venous system were found. Of all the insular veins, the precentral insular vein was the one that most commonly connected to the superficial sylvian vein.
Introduction
The insula is a roughly triangular area, located deep to the frontal, parietal, and temporal opercula in the floor of the sylvian fissure. Complete exposure of the insula requires that the sylvian fissure be opened widely. The technical complexity involved in opening the sylvian fissure for insular exposure and the vital pathways coursing deep to the insula make surgery for vascular and neoplastic lesions in this area hazardous.3–5,9,16,18,23,31,43,45,46 There have been numerous reports on the anatomy of the sylvian fissure and insula.1,2,10,12,28–30,32,39,40 Nonetheless, a detailed description of the topographic anatomy, arterial supply, and venous drainage of the area is still lacking. The purpose of this study was to define clearly the neural features, arterial supply, and venous drainage of the insula and its overlapping opercula. An understanding of these relationships should facilitate a safer, more precise surgical exploration of the region.
Materials and Methods
The sylvian fissure and insula were examined using x3 to x40 magnification in 43 cadaveric cerebral hemispheres in which the arteries and veins had been perfused with colored silicone. A pterional craniotomy was performed, and the entire sylvian fissure was exposed. The surface variations in the sulci and gyri along the sylvian fissure and the pattern of drainage of the SSV were examined prior to dissection. The sylvian fissure was opened widely by using microsurgical methods, and the variations in the opercular and insular sulci and gyri as well as in the rami of the sylvian fissure were noted. The MCA bifurcation was identified, and the insular area supplied by each branch was described. Each insular vein’s pattern of drainage and its connection to the SSV and deep MCV were identified. The relationship between surgically important landmarks, such as the LSAs, limen insulae, and insular pole and apex, were detailed.

FIG. 1. A-F. Photographs of cadaveric brains obtained during stepwise dissection of right cerebral hemisphere. A: The natural upward retraction of the apex of the pars triangularis creates the largest natural opening in the sylvian fissure. The posterior ramus, the longest ramus of the sylvian fissure, extends backward and upward to its termination in the inferior parietal lobule, where the supramarginal gyrus wraps around its posterior end. The Heschl gyrus, the most anterior of the transverse temporal gyri, extends obliquely backward and medially across the temporal operculum from the cortical surface. The frontoorbital ramus of the sylvian fissure crosses the basal surface of the frontal lobe medial to the pars orbitalis. B: The lower and upper portions of the frontal and temporal opercula, respectively, have been removed to show the relationship between opercular and insular structures. The central insular sulcus courses superficial to, and almost parallel with, the central sulcus on the convexity. In this hemisphere, the frontoorbital ramus (red dotted arrow) is differentiated from a superficial orbital sulcus (white arrow)— which is located on the orbital surface of the frontal lobe between the anterior horizontal and frontoorbital rami of the sylvian fissure—by the fact that the deep end of the frontoorbital ramus opens into the sylvian fissure at the level of the anterior limiting sulcus, but the orbital sulcus above it does not open into the sylvian fissure. The mean length of the frontoorbital ramus of the sylvian fissure is 10 mm. Opening the frontoorbital ramus, when present, provides access to the most anteroinferior portion of the insula. C: The remaining part of the frontoparietal operculum has been removed. The anterior limiting sulcus is directed upward and anterior, to form the anterior border of the insula. The insular apex is the most prominent laterally projecting point on the insular convexity. D: Enlarged view of the insula. The middle short gyrus is the widest of the short gyri in this specimen.

FIG. 1. G-L. Photographs of cadaveric brains featuring stepwise dissection of right cerebral hemisphere. G: The temporal operculum has been retracted to expose the inferior part of the insula. The pars triangularis is small and its tip is retracted upward away from the fissure, resulting in the formation of a wide space or a wide common stem (red dotted arrow) between the partes opercularis and orbitalis, from which the anterior horizontal and ascending rami arise. H: The frontoparietal and temporal opercula have been retracted to expose the insula. The inferior limiting sulcus is positioned below the long gyri of the insula and separates the insula from the sylvian surface of the temporal lobe. The central sulcus, the deepest insular sulcus, separates the insula into larger anterior and smaller posterior portions. I: Lateral view of the same hemisphere. The anterior and middle short gyri are separated by the short insular sulcus, and the middle and posterior short gyri are separated by the precentral insular sulcus. The long gyri arise as a single gyrus near the limen insulae and bifurcate posteriorly into anterior and posterior long gyri, which are separated by the long insular sulcus. The transverse gyrus connects the anterior part of the insula to the gyri on the orbital surface. The insular pole, located at the anteroinferior edge of the insula between the anterior limiting sulcus and the central insular sulcus, is approximately 2 cm wide (green area). The insular pole is located below the insular apex, the most laterally prominent point of the insula. J: The frontoparietal operculum was removed using an axial cut at the level of the superior limiting sulcus, and the temporal operculum was removed using a sagittal cut parallel to the lateral surface of the temporal lobe. The most anterior of the transverse temporal gyri, the Heschl gyrus, extends backward from the cortical surface to the junction of the superior and inferior limiting sulci. K: The sagittal cut has been extended to the medial edge of the inferior limiting sulcus to show the relationship between the temporal horn and the insula. The Heschl gyrus is located above the roof of the temporal horn and the body of the hippocampus. The vertical distance from the inferior limiting sulcus to the posterior edge of the head of the hippocampus is approximately 1 cm. L: Lateral view of another right insula. The frontoparietal and temporal opercula have been removed to expose the insula. The central insular sulcus, the deepest of the insular sulci, arises from the limen area and extends posterosuperiorly to reach the superior limiting sulcus. The anterior part of the insula consists of three gyri—the anterior, middle, and posterior short gyri. These are separated by the short insular and precentral insular sulci and fuse with each other at the insular pole. The long gyri arise as a single gyrus below the insular apex, near the limen insulae, and bifurcate posteriorly into two gyri. Incising along the inferior limiting sulcus has exposed the temporal horn of the lateral ventricle. The choroid plexus has been elevated to expose the head and body of the hippocampus. The inferior choroidal point, the lower end of the choroidal fissure, is located just behind the head of the hippocampus at a distance of approximately 1 cm from the inferior limiting sulcus. The anterior choroidal artery enters the ventricle at the inferior choroidal point. A. = arteries or artery; Acc. = accessory; ALG = anterior long gyrus; Ant. = anterior; Asc. = ascending; ASG = anterior short gyrus; Call. = callosum; Caps = capsule; Cent. = central; Ch. = choroid or choroidal; Coll. = collateral; Corp. = corpus; Emin. = eminence; For. = foramen; Fr. = fronto or frontal; Gyr. = gyrus; Hippo. = hippocampus; Hor. = horizontal; Inf. = inferior; Ins. = insulae or insular; Lent. = lentiform; Lim. = limiting; Mid. = middle; MSG = middle short gyrus; Nucl. = nucleus; Oper. = opercularis or operculum; Orb. = orbital or orbitalis; Pell. = pellucidum; Plex. = plexus; PLG = posterior long gyrus; Post. = posterior; Postcent. = postcentral; Precent. = precentral; PSG = posterior short gyrus; Ram. = ramus; Sept. = septum; Sh. = short; Sul. = sulcus; Sup. = superficial or superior; Supramar. = supramarginal; Temp. = temporal; Tr. = tract or transverse; Tri. = triangularis.
Results
Topographical Anatomy
Sylvian Fissure
The sylvian fissure is the most distinct landmark on the lateral surface of the cerebrum.20,33,35 It provides passage to the MCA and its branches and provides a surgical gateway connecting the cerebral surface to the anterior part of the basal surface of the brain and skull base. The superficial part of the sylvian fissure is composed of a stem and several rami. This stem begins medially at the anterior clinoid process, extends laterally behind the sphenoid ridge, and ends at the convexity by dividing into the anterior horizontal, anterior ascending, and posterior rami (Figs. 1 and 2). The posterior ramus, the longest of the sylvian fissure, extends backward and upward from the region of the pterion to its termination in the inferior parietal lobule, where the supramarginal gyrus wraps around its posterior end. The shorter anterior horizontal and anterior ascending rami, present in all hemispheres, have nearly identical lengths and divide the inferior frontal gyrus, from anterior to posterior, into the partes orbitalis, triangularis, and opercularis (Figs. 1 and 2 and Table 1). The anterior horizontal ramus separates the partes orbitalis and triangularis; the anterior ascending ramus separates the partes triangularis and opercularis. Opening the anterior horizontal and ascending rami provides access to the upper anterior portion of the insula; opening the posterior ramus and adjacent stem provides access to the remaining portion of the insula.
An additional ramus, the frontoorbital ramus, was found in 27 of 43 cadaveric hemispheres. This ramus, when present, crossed the basal surface of the frontal lobe below the anterior horizontal ramus and pars orbitalis (Fig. 1). The deep portion of the posterior end of the frontoorbital ramus was positioned approximately midway between the upper and lower ends of the anterior limiting sulcus (Fig. 2). The ramus separated the pars orbitalis from the posterior orbital gyrus. A short shallow sulcus may cross the lateral aspect of the frontal lobe below the anterior horizontal ramus, but it is not considered to be a ramus of the sylvian fissure because it does not reach the insula in its depths. This contrasts with the frontoorbital ramus, which crosses the basal surface of the frontal lobe below the anterior horizontal ramus and opens into the insula, as do the anterior horizontal and ascending rami (Fig. 1).
The deep portion of the sylvian fissure is divided into sphenoidal and operculoinsular compartments.6,20,22 The sphenoidal compartment lies proximal to the limen area behind the sphenoidal ridge, and the operculoinsular compartment is positioned deep to the superficial rami of the sylvian fissure. This latter compartment is formed by two narrow clefts: opercular and insular. The opercular cleft is situated between the opposing lips of the frontoparietal operculum above and the temporal operculum below. The insular cleft, situated between the insula and the medial surface of the opercula, has two limbs. The superior limb of the insular cleft is located between the insula and the medial surface of the frontoparietal operculum; the inferior limb is situated between the insula and the medial surface of the temporal operculum.
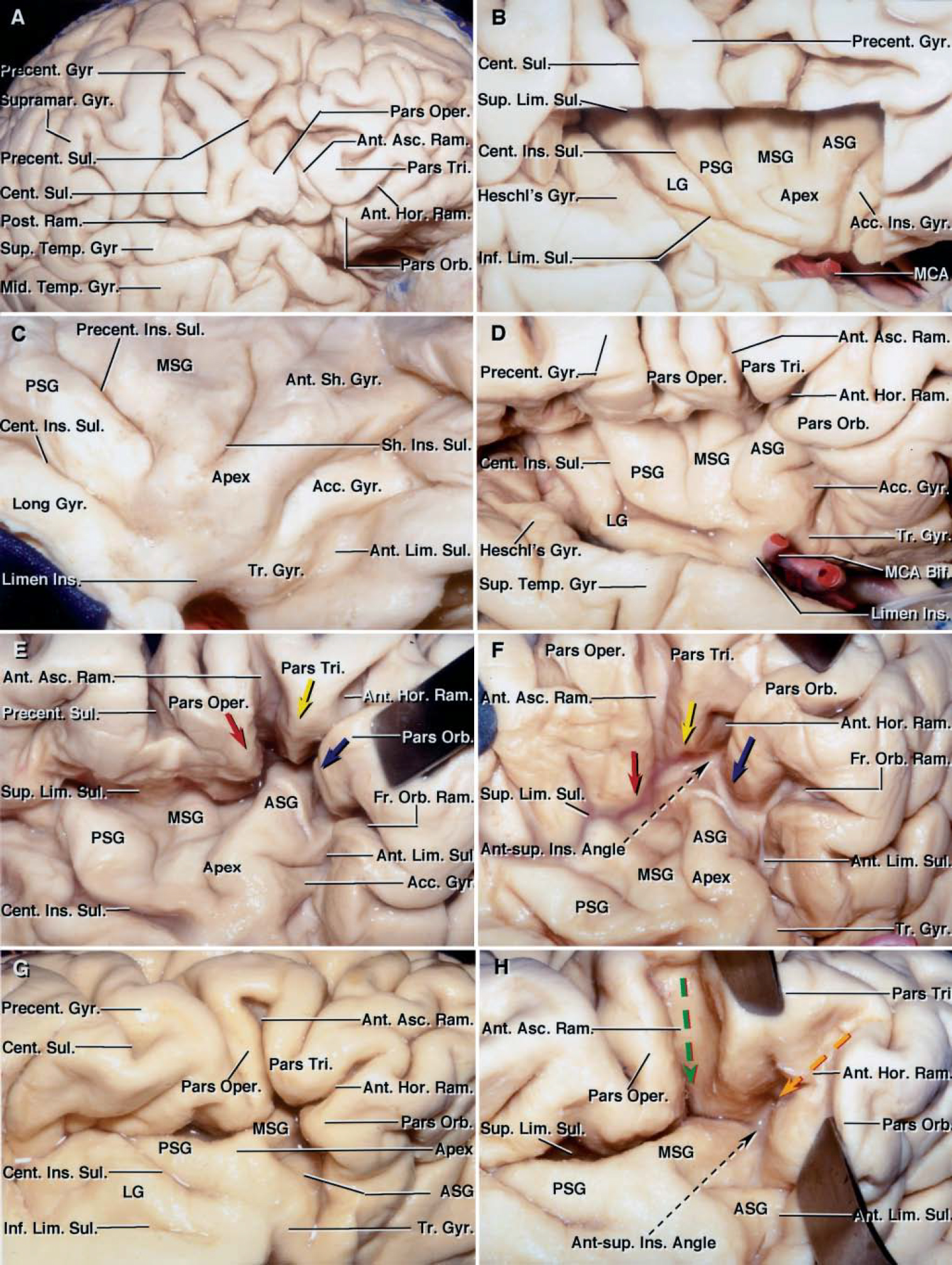
FIG. 2. Photographs of cadaveric brains obtained during stepwise dissection of right cerebral hemisphere. A: The inferior frontal gyrus is composed of the partes orbitalis, triangularis, and opercularis. A gyral bridge connects the lower end of the pre- and postcentral gyri so that the central sulcus does not open into the sylvian fissure. The posterior ramus of the sylvian fissure turns upward into the supramarginal gyrus at its posterior end. B: The frontoparietal operculum has been removed to expose the insula and show the relationship between the central sulcus on the convexity and the central insular sulcus. The inferior end of the central sulcus on the convexity is located less than 5 mm posterior to the central insular sulcus. The Heschl gyrus, the most anterior transverse temporal gyrus, extends obliquely backward and medially from the cortical surface and intersects the posterosuperior angle of the insula. The superior limiting sulcus is oriented horizontally and separates the upper border of the insula from the sylvian surface of the frontal and parietal lobes. The central insular sulcus divides the insula into a larger anterior portion and a smaller posterior portion. Typically, there are three anteriorly placed short gyri that form a radiating pattern converging at the anteroinferior end of the insula. In this hemisphere, there is a fourth short gyrus, the accessory insular gyrus, positioned in front of the other three. The lower end of the accessory gyrus blends into the transverse gyrus, which in turn blends into the orbital gyri. The posterior portion of the insula behind the central insular sulcus consists of a single long gyrus in this hemisphere. C: Anterolateral view of the insula in another right cerebral hemisphere. The frontal operculum has been retracted to expose the most anterior portion of the insula. The anterior, middle, and posterior short gyri form a radiating pattern that converges at the anteroinferior end of the insula. The insular apex is the highest or most prominent laterally projecting site on the insular cortex. The insular pole, the anteroinferior edge of the insula below the apex, is located between the anterior limiting sulcus and the central insular sulcus, and blends inferomedially into the gyri on the orbital surface of the frontal lobe. The deep end of the anterior margin of the pole is joined to the posterior orbital gyri by the transverse insular gyrus. The accessory insular gyrus is located superior to, and blends into, the transverse gyrus. The anterior limiting sulcus extends obliquely upward and forward, deep to the frontal operculum. Photographs obtained during stepwise dissection of right hemisphere (D–F) display the relationships between the anterior horizontal, ascending, and frontoorbital rami of the sylvian fissure, and the insular sulci and gyri. D: The frontoparietal and temporal opercula have been retracted to expose the insula. The most consistent rami of the sylvian fissure are the anterior horizontal, anterior ascending, and posterior rami. The anterior ascending and anterior horizontal rami divide the inferior frontal gyrus, from anterior to posterior, into the partes orbitalis, triangularis, and opercularis. The MCA bifurcation is located at the level of the limen insulae in this hemisphere. E: The inferior frontal gyrus has been retracted anterosuperiorly to open partially the anterior horizontal and ascending rami of the sylvian fissure. An additional anterior ramus of the sylvian fissure, called the “frontoorbital ramus,” arises below the anterior horizontal ramus, medial to the pars orbitalis, and crosses the basal surface of the frontal lobe. Opercular landmarks can be used to localize deeper structures in the insular area. The apices of the partes triangularis (yellow arrow), orbitalis (blue arrow), and opercularis (red arrow) lie superficial to the anterior short gyrus and the adjacent short sulci. The free surface of the three parts of the inferior frontal gyrus, if followed to the depths, ends at the limiting sulci. F: Inferolateral view directed along the deep side of the frontal operculum. The anterior ascending, anterior horizontal, and frontoorbital rami have been opened to show the relationship between the opercular landmarks and the insula. Opening the anterior horizontal, anterior ascending, and frontoorbital rami plus the adjacent portion of the posterior ramus of the fissure provides access to the upper anterior portion of the insula. Opening the anterior ascending ramus exposes the superior limiting sulcus. Opening the anterior horizontal ramus exposes the anterosuperior angle of the insula formed by the junction of the superior and inferior limiting sulci. Opening the frontoorbital ramus of the sylvian fissure exposes the anterior limiting sulcus. The upper edge of the pars orbitalis lies superficial to the upper portion of the anterior short gyrus and the adjacent portion of the superior limiting sulcus (blue arrow). The upper end of the pars triangularis is positioned superficial to the upper portion of the anterior short gyrus and the adjacent portion of the superior limiting sulcus (yellow arrow). The upper end of the pars opercularis lies superficial to the superior portion of the middle short gyrus and the adjacent portion of the superior limiting sulcus (red arrow). Photographs obtained during stepwise dissection of another right hemisphere (G and H), showing the relationships between the anterior ascending and anterior horizontal rami, and the insula. G: The anterior horizontal ramus separates the partes orbitalis and triangularis; the anterior ascending ramus separates the partes triangularis and opercularis. H: Opening the anterior horizontal ramus exposes the anterosuperior insular angle and the adjacent portion of the middle short gyrus (orange arrow) in this specimen. Opening the anterior ascending ramus exposes the superior limiting sulcus (green arrow) and the adjacent portion of the middle short gyrus. The superior limiting sulcus of the insula lies in the depths of the upper end of the anterior ascending ramus (green arrow). The anterosuperior insular angle lies in the depth of the anterior end of the anterior horizontal ramus (orange arrow). Ant-sup. = anterosuperior; Bif. = bifurcation; LG = long gyrus. See legend to Fig. 1 for definitions of additional abbreviations.

Table 1. Sylvian fissure and opercular measurements in 43 cadaveric hemispheres.

Table 2. Insular landmark measurements in 43 cadaveric hemispheres.
Insula
The insular cortex faces laterally and forms the medial wall of the operculoinsular compartment. It is encircled and separated from the frontal, parietal, and temporal opercula by a shallow sulcus, the circular sulcus.33,35 This sulcus is frequently referred to as the “limiting sulcus” because it surrounds the periphery of the insula. It is more triangular than circular in shape because of the pyramidal configuration of the insula (Fig. 2). The sulcus has three parts: anterior, superior, and inferior. Its anterior border, referred to as the “anterior limiting sulcus,” from its lower end is directed upward and forward, deep to the pars orbitalis of the frontal operculum. The superior border, or the superior limiting sulcus, is oriented horizontally. It extends beneath the frontoparietal operculum from the upper end of the anterior limiting sulcus at the anterosuperior edge of the insula to the posterior end of the inferior limiting sulcus. Beneath the temporal operculum at the lower edge of the insula lies the inferior limiting sulcus. The longest limiting sulcus was the superior; the shortest, the anterior (Figs. 1 and 2).
The insular cortex is composed of three anteriorly placed short gyri and two posteriorly placed long gyri, which are separated by the central insular sulcus. Two anterior sulci separate the three short gyri, and a single long sulcus separates the two long gyri (Fig. 1). The sulci and gyri of the insula form a radial pattern extending posteriorly and superiorly from the anteroinferior portion of the insula that borders the limen insulae (Figs. 1 and 2). The central insular sulcus, the deepest of the insular sulci, was present in all hemispheres. It extended posteriorly and superiorly from the limen insulae in approximately 70% of the hemispheres; in the remaining hemispheres, it began between 3.4 to 12.3 mm behind the limen insulae. It reached the superior limiting sulcus in all hemispheres. Furthermore, the central insular sulcus coursed almost parallel to the central (rolandic) sulcus of the hemisphere. The central sulcus extended around the opercular lip and was continuous with the central insular sulcus in nine hemispheres. In the remaining hemispheres, in which the central sulcus did not blend into the central insular sulcus, the inferior end of the rolandic sulcus was positioned from 5.9 mm posterior to 5 mm anterior to the central insular sulcus.
The central insular sulcus divided the insula into a larger anterior portion formed by the short gyri and a smaller posterior portion formed by the long gyri. The anterior portion of the insula, that part located in front of the central insular sulcus, was composed of three gyri in 41 hemispheres and four gyri in two hemispheres. The widest of the short gyri was the anterior one in 27 hemispheres, the middle one in four, and the posterior one in 12. The short gyri of the insula were separated by two sulci; the short insular sulcus separated the anterior and middle short gyri, and the precentral insular sulcus separated the middle and posterior short gyri (Figs. 1 and 2).
Two important points situated on the insula include the pole and the apex. The insular pole is located at the anteroinferior edge of the insula, where the short gyri converge to form a rounded area lateral to the limen (Figs. 1 and 3 and Table 2). The insular apex is the highest and most prominent laterally projecting area on the insular convexity. It is located above and behind the pole on the short gyri, usually on the middle short gyrus (Fig. 3 and Table 2).
The deep end of the anterior margin of the pole is connected to the posterior orbital gyri of the frontal lobe by the transverse gyrus of the insula (Figs. 1 and 2). The transverse gyrus, the shortest insular gyrus, was present in all hemispheres. It is a narrow gyrus, extending medially from the pole and blending into the posterior orbital gyrus. Another small gyrus, the accessory insular gyrus, was present in 26 hemispheres (60%). It was located superior to the transverse insular gyrus just behind the lower two thirds of the anterior limiting sulcus and blended below into the lateral end of the transverse gyrus. The transverse gyrus blended superiorly into the anterior limiting sulcus if the accessory gyrus was absent (Fig. 1).
The long gyri are located in the posterior and inferior portions of the insula, behind the central insular sulcus. The posterior portion of the insula was formed by one long gyrus in three hemispheres and two long gyri in 40 hemispheres. The long gyri most frequently arose from below the insular apex, near the limen insulae, as a single gyrus that bifurcated posteriorly into anterior and posterior long gyri, which were separated by the long insular sulcus (Fig. 1). The anterior long gyrus was the widest of the long gyri in the majority of cadaveric hemispheres. The long insular sulcus, when present, extended downward and forward from the superior limiting sulcus and terminated near the limen in most hemispheres.
The limen insulae is a slightly raised, arched ridge located at the junction of the sphenoidal and operculoinsular compartments of the sylvian fissure and extends from the temporal pole to the orbital surface of the frontal lobe.36 It overlies the uncinate fasciculus and is covered by a thin layer of gray matter. Its width, measured from its lateral edge at the anterior end of the long gyrus, where it fuses with the temporal pole, to its medial edge at the middle of the posterior orbital gyrus, was a mean 21.36 mm (range 16.37– 29.84 mm; Figs. 1 and 2 and Table 2).
The anterior perforated substance lies just medial to the limen insulae and serves as an important surgical landmark. In this study, the point of entrance of the most lateral LSA was considered to be the lateral limit of the anterior perforated substance. The mean distance from the point of entrance of the most lateral LSA into the anterior perforated substance to the medial border of the limen insulae was 15.3 mm (range 9.77 to 22.61 mm). There was a shallow recess, referred to as the “limen recess,” between the medial border of the limen insulae and the point of entrance of the most lateral LSA in all hemispheres; this recess was devoid of important perforating arteries (Fig. 4 and Table 2).
The most lateral LSA originated from the M1 segment prior to bifurcation in 16 of 43 hemispheres, from the inferior trunk in 14, and from the superior trunk in nine. The distance between the origin of the most lateral LSA and the insular apex was less than 23 mm in all hemispheres (Fig. 3 and Table 1).
Sylvian Fissure–Insular Relationships
Opening specific portions of the sylvian fissure can expose different parts of the insula. For example, opening the posterior ramus below the apex of the pars triangularis exposed the anterior and middle short gyri and the anteroinferior portion of the insula. Opening the anterior horizontal ramus exposed the upper portion of the anterior short gyrus and the adjacent junction of the superior and anterior limiting sulci in 44% of the hemispheres, the superior limiting sulcus alone in 35%, and the anterior limiting sulcus alone in the remaining 21% (Fig. 2). Opening the anterior ascending ramus exposed the anterior short gyrus and the adjacent portion of the superior limiting sulcus in the majority of hemispheres. Opening the frontoorbital ramus, present in nearly 65% of hemispheres, provided access to the most anteroinferior portion of the insula, including the anterior limiting sulcus.
The posterior, anterior horizontal, and anterior ascending rami converge below the apex of the pars triangularis (Figs. 1 and 2 and Table 1). The natural upward retraction of the apex of the pars triangularis away from the fissure provides the largest surface opening in the sylvian fissure and is a site naturally suitable to begin the opening of the sylvian fissure. In eight of 43 hemispheres, the pars triangularis was small in size, resulting in the formation of a common stem from which the anterior horizontal and anterior ascending rami arose.
Opercular landmarks at the cerebral surface can be used to localize deeper structures in the insular area. Following the opercular surface of pars orbitalis to its deep edge exposed the upper part of the anterior short gyrus and adjacent part of the anterior limiting sulcus (Fig. 2). Following the cortical surface of the pars opercularis exposed the superior portion of the anterior or middle short gyri and the adjacent portion of the superior limiting sulcus. Following the upper edge of the pars triangularis exposed the upper portion of the anterior short gyrus in most hemispheres, but had a more variable relationship to the limiting sulcus. The upper edge of the pars triangularis was positioned superficial to the superior limiting sulcus in 39 of 43 hemispheres, to the anterior limiting sulcus in two, and to the junction of the anterior and superior limiting sulci in two. The inferior limiting sulcus was located medial to the superior temporal sulcus. The limen insulae, the site of the MCA bifurcation and the origin of the trunks, was located medial to the temporal operculum; the site of the main branching of the MCA trunks was located medial to the frontal operculum.
The distance from the cortical surface, measured along the surface of the opercular lips to the superior and inferior limiting sulci of the insula, varied from anterior to posterior. Anteriorly, the distance from the cortical surface along the upper opercular lip to the superior limiting sulcus was greater than that along the lower opercular lip to the inferior limiting sulcus. Posteriorly, the distances were nearly identical, but in some cases the distance to the inferior limiting sulcus was greater than the distance to the superior limiting sulcus. The distance along the cortical surface was approximately 18 mm from the lateral apex of the pars triangularis to the superior limiting sulcus of the insula (Fig. 3 and Table 1).
The supramarginal gyrus surrounds the upturned posterior end of the sylvian fissure (Figs. 1 and 2). This gyrus lies superficial to the junction of the superior and inferior limiting sulci at the posterior end of the insula. The mean depth of the junction of the superior and inferior limiting sulci was 25 mm from the convexity at the posterior end of the sylvian fissure (Fig. 3 and Table 1).
The superior temporal gyrus forms the superficial portion of the temporal operculum and is continuous with the transverse temporal gyri on the upper surface of the temporal lobe (Figs. 1 and 2). The transverse gyri extend obliquely backward and medially from the cortical surface toward the posterosuperior angle of the insula. The length of the most anterior transverse temporal gyrus, the Heschl gyrus, was a mean 34 mm (range 22.25–40.78 mm) from the cortical surface to the junction of the superior and inferior limiting sulci.

FIG. 3. Drawings of the lateral view of the left opercular and insular areas. A: The opercular and insular areas are labeled. B: Distances between the points in this figure are listed in Table 1. Points A, C, and E were located at the apices of the partes orbitalis, triangularis, and opercularis, respectively. The measurements from A to B, C to D, and E to F are the depths from the cortical surface along the opercular lips of the inferior frontal gyrus to the anterior and superior limiting sulci. Point I was located at the proximal end of the Heschl gyrus on the cortical surface of the superior temporal gyrus, and Point H was situated at the posterior end of the Heschl gyrus (the junction of the superior and inferior limiting sulci). The measurement from G to H was the distance between the supramarginal gyrus and the posterior end of the insula. Point J was located at the insular apex, the highest or the most prominent laterally projecting point on the insular convexity. Point K was situated at the origin of the most lateral LSA from the MCA. C: Enlarged view of the insula. The distances between the points in this figure are featured in Table 2. The points are located as follows: A, inferior end of central insular sulcus; B, midpoint of anterior limiting sulcus; C, inferior limit of limen insulae; D and F, junctions of anterior and superior, and superior and inferior limiting sulci, respectively; E and H, midpoints of superior and inferior limiting sulci, respectively; and G, apex of insula. See legend to Fig. 1 for definitions of abbreviations.

FIG. 4. Photographs featuring the limen recess. A: Anteroinferior view of the anterior perforated substance and the limen recess in left hemisphere. The frontoparietal and temporal opercula have been retracted to expose the anteroinferior part of the insula. The limen recess is the area between the medial edge of the limen insulae and the point at which the most lateral LSA enters the anterior perforated substance. It has a mean width of 15 mm. No LSAs enter this area. The anterior perforated substance is usually considered to extend lateral to the limen insulae; however, the limen recess is devoid of perforating branches and forms the lateral limit of the anterior perforated substance. The accessory gyrus, located in front of the anterior short gyrus, blends below into the transverse gyrus. The transverse gyrus, the shortest of all insular gyri, connects the anterior edge of the insula to the posterior orbital gyrus. B: Enlarged inferolateral view of another left cerebral hemisphere. The frontoparietal operculum has been removed, and the temporal operculum has been retracted to expose the insula. The limen insulae extends from the anterior end of the long gyri, where it fuses with the temporal pole, to the posterior orbital gyrus. The limen recess lies between the point of entry of the most lateral LSA into the anterior perforated substance and limen insulae. The transverse gyrus is positioned between the insular pole and the posterior orbital gyri. C: Inferior view of left cerebral hemisphere. The anterior perforated substance forms the roof of the sphenoidal compartment of the sylvian fissure. The right temporal lobe has been removed down to the level of the temporal stem. The anterior perforated substance has a salt-and-pepper appearance, created by small openings through which the perforating arteries and veins penetrate the hemisphere. The limen recess, located between the point at which the most lateral LSA enters the anterior perforated substance and the limen insulae, is devoid of openings for perforating arteries. APS = anterior perforated substance; CN = cranial nerve; Lat. = lateral; Med. = medial; Olf. = olfactory; Pit. = pituitary; Rec. = recurrent. See legend to Fig. 1 for definitions of additional abbreviations.
Arterial Relationships
The MCA
The MCA, the largest and most complex of the three cerebral arteries, provides the sole supply to the insula. The MCA is divided into four segments: M1 (sphenoidal), M2 (insular), M3 (opercular), and M4 (cortical) (Fig. 5).6,8,22 The M1, or sphenoidal, segment begins at the origin of the MCA and extends laterally within the depths of the sylvian fissure. The M1 segment ends and the M2 segment begins at the site of a 90˚ turn, the genu, usually located at the level of, or immediately distal to, the limen insulae. The M2 segment includes the trunks that lie on and supply the insula. The M2 segment ends and the M3 segment begins at the limiting sulci along the periphery of the insula. The M3 segment courses around the opercular lips and ends at the cortical surface of the sylvian fissure. The M4 segment is composed of the branches to the cerebral convexity (Figs. 5 and 6).
The main trunk of the M1 segment ended in a bifurcation in 38 hemispheres and a trifurcation in the remaining five hemispheres. The postbifurcation trunks of the M1 segment ran nearly parallel to each other, diverging only minimally prior to reaching the genu (Fig. 5). The postbifurcation trunks underwent extensive branching distal to the genu, near the insular pole, at the level of the apex of the insula and coursed either posterosuperiorly or posteroinferiorly to reach the limiting sulci (Fig. 6). The M2 segment, as it crossed the insula before reaching the superior and inferior limiting sulci, gave rise to a mean of eight branches (range five–11 branches). The branches arising from the postbifurcation trunks that give rise to two or more cortical arteries are referred to as “stem arteries.” The majority of the stem arteries divided into their individual cortical branches at or before reaching the limiting sulci. Of the 329 branches arising from the postbifurcation trunk and the early branches in 43 hemispheres, 214 were arteries supplying a single cortical area and 115 were stem arteries giving rise to two or more cortical arteries. Of the 115 stem arteries, 98 produced two and 17 generated three or more cortical arteries.

FIG. 5. A: Photograph obtained during a left pterional exposure. Inset features the position of the head and the scalp incision. The M1 segment begins at the origin of the MCA, extends laterally within the depths of the sylvian fissure, crossing below the anterior perforated substance, and ends at the site of a 90° turn, the genu, located at the level of the limen insulae. The postbifurcation trunks of the M1 segment ran nearly parallel to each other, diverging only minimally prior to reaching the genu. Photographs featuring stepwise dissections of left cerebral hemisphere (B and C). B: Anterior view. The coronal cross-section extends through the foramen of Monro. The ICA ascends lateral to the optic nerve. The M1 segment begins at the origin of the MCA, extends laterally below the anterior perforated substance, and ends at the genu, located at the level of the limen insulae. It gives rise to the LSAs. The M2 segment begins at the genu, where the MCA trunks pass the limen insulae, and ends at the limiting sulci along the periphery of the insula. The M3 segment courses around the opercular lips. The sylvian point is located at the point at which the most posterior branch of the M2 segment turns away from the insular surface toward the lateral convexity near or at the junction of the superior and inferior limiting sulci. C: Anterosuperior view of the same hemisphere. The coronal cut has been extended to the level of the lateral geniculate body, and the temporal horn has been exposed through an additional axial cut. The M2 segment crosses the insula above and lateral to the temporal horn. The M3 branches turn laterally away from the insula and cross the transverse temporal gyri to reach the cortical surface. The most anterior transverse temporal gyrus forms the Heschl gyrus. D: Photograph of another specimen of the left cerebral hemisphere, left pterional exposure. The sphenoidal compartment of the sylvian fissure sits behind the sphenoidal ridge in the area proximal to the limen insulae. The M1 segment gives rise to early frontal and temporal branches and bifurcates at the level of the limen insulae. The LSAs arise from both of the early branches. A1 = A1 segment of the ACA; Bas. = basilar; Br. = branch; Cer. = cerebral; Clin. = clinoid; Com. = communicating; Gen. = geniculate; Glob. = globus; Int. = internal; Lat. = lateral; M1 = M1 segment of the MCA; M2 = M2 segment of the MCA; M3 = M3 segment of the MCA; P. = point or posterior; Pall. = pallidus; S.C.A. = superior cerebellar artery; Tent. = tentorium; Tr. = tract, transverse, or trunk; V. = vein. See legends to Figs. 1, 2, and 4 for definitions of additional abbreviations.

FIG. 6. Photographs obtained during stepwise dissections of the right cerebral hemisphere (A–C), demonstrating the insular and opercular branches of the MCA. A: Lateral view of the hemisphere showing the gyri and sulci bordering the sylvian fissure, which are supplied by the cortical branches of the MCA. The cortical arteries include the orbitofrontal, prefrontal, precentral, central, anterior parietal, posterior parietal, angular, temporooccipital, posterior temporal, middle temporal, anterior temporal, and temporopolar arteries. The vein of Trolard runs between the SSV and the superior sagittal sinus. The SSV also has a large anastomosis with the vein of Labbé. B: The frontoparietal operculum has been removed while preserving the branches that form the M2 and M3 segments. The cortical arteries can be followed from their origin along the insula to the cortex. The M1 bifurcates proximal to the genu near the limen insulae. The early temporal branch, arising from the prebifurcation M1 segment, gives rise to the temporopolar, anterior temporal, middle temporal, posterior temporal, and temporooccipital arteries. The superior trunk gives rise to orbitofrontal, prefrontal, and precentral arteries along its course around the pole of the insula. The inferior trunk gives rise to the central, anterior and posterior parietal, and angular arteries. All of the cortical branches arising from the early branch, except for the temporopolar artery, course along the posterior long gyrus and inferior limiting sulcus, and contribute to the supply of these insular areas. C: Enlarged view of the insula. At the pole of the insula, the superior trunk gives rise to its first cortical branch, the orbitofrontal artery, prior to reaching the insular apex. The orbitofrontal artery courses along the most anteroinferior part of the insula and contributes to the supply of the anterior short gyrus. The prefrontal and precentral arteries arise from a common stem artery (black arrow) near the level of the insular apex. The prefrontal artery courses along and supplies the middle short gyrus en route to the anterosuperior angle of the insula. The precentral artery courses near the precentral insular sulcus. The central artery, the first cortical branch of the inferior trunk, supplies the central insular sulcus and the posterior short gyrus. The first cortical branch from the inferior trunk, the central artery, arises farther away from the MCA bifurcation than the first cortical branch of the superior trunk. The central insular sulcus does not receive a contribution from the superior trunk in this hemisphere. The anterior long gyrus is supplied by a stem artery (red arrow) from the inferior trunk that eventually gives rise to the angular and the anterior and posterior parietal arteries. Photographs obtained during stepwise dissections of another right cerebral hemisphere (D–F). D: Lateral view showing the cortical territory supplied by the MCA, which includes the majority of the lateral surface of the hemisphere, the lateral part of the orbital surface of the frontal lobe, the temporal pole, and the lateral part of the inferior surface of the temporal lobe. This territory is supplied by the following cortical MCA branches: the orbitofrontal, prefrontal, precentral, central, anterior parietal, posterior parietal, angular, temporooccipital, posterior temporal, middle temporal, anterior temporal, and temporopolar arteries. E: The frontoparietal and temporal opercula have been removed while preserving the branches forming the M2 and M3 segments. The M1 segment bifurcates proximal to the genu near the limen insulae. The first cortical branch of the superior trunk is an orbitofrontal artery that supplies the accessory gyrus. The superior trunk gives rise to the posterior parietal artery on the anteroinferior portion of the insula (black arrow) prior to reaching the insular apex. The posterior parietal artery courses along the central insular sulcus. The central and anterior parietal arteries arise at the level of the insular apex, from a common stem (red arrow) of the superior trunk that courses along the posterior short gyrus. The prefrontal and precentral arteries arise from a common stem artery above the insular apex. The prefrontal artery sends perforating arteries to the anterior short gyrus and crosses the anterosuperior angle of the insula. The inferior trunk gives rise to the angular and temporooccipital arteries. The posterior and the middle and anterior temporal arteries arise from the early temporal branch. F: Enlarged view of the same hemisphere. The orbitofrontal artery, the first cortical branch of the superior trunk, sends branches to the accessory gyrus. The posterior parietal artery arises from the superior trunk at the level of the insular pole below the apex, courses along the central insular sulcus, and sends perforating vessels to the posterior short and anterior long gyri. The stem artery (red arrow), which gives rise to the central and anterior parietal arteries, arises from the superior trunk at the level of the insular apex, supplies the posterior short gyrus and precentral sulcus, and gives rise to a large insular perforating artery near the superior portion of the insula. The prefrontal and precentral arteries arise from a common stem artery above the insular apex and send perforating arteries to the anterior and middle short gyri, respectively. The angular and temporooccipital arteries arise from a stem artery (green arrow) that originates from the inferior trunk. The stem artery supplies the posterior long gyrus and inferior limiting sulcus. Photographs obtained during the stepwise dissection of a right cerebral hemisphere (G and H), demonstrating the insular, opercular, and cortical segments of the MCA. G: Lateral view of the cortical surface bordering the sylvian fissure. The anterior horizontal ramus separates the partes orbitalis and triangularis, and the anterior ascending ramus separates the partes triangularis and opercularis. The lower end of the precentral gyrus is located behind the pars opercularis. H: The frontal and temporal opercula have been retracted to expose the insular, opercular, and cortical course of the prefontal, precentral, central, anterior and posterior parietal, and angular arteries. The M1 segment ends and the M2 segment begins at the site of a 90° turn, the genu, located distal to the MCA bifurcation. The M2 segment includes the trunks that lie on and supply the insula. The M1 segment of the MCA bifurcates at the level of the limen insulae. The superior trunk gives rise to the orbitofrontal, prefrontal, and precentral arteries. At the pole of the insula, the orbitofrontal artery arises from the initial centimeter of the superior trunk, proximal to the level of the insular apex, and courses anteroinferiorly to supply the orbital gyri of the frontal lobe. The prefrontal and precentral arteries arise from a common stem artery at the level of the insular apex. The prefrontal artery courses along the middle short gyrus to reach the junction of the superior and inferior limiting sulci and sends branches to the anterior short gyrus. The precentral artery courses on the precentral sulcus. The initial portion of the inferior trunk supplies the limen insulae and gives rise to the central artery as its first cortical branch. The first cortical branch from the inferior trunk, the central artery, arises farther from the MCA bifurcation than the first cortical branch of the superior trunk, the orbitofrontal artery. The central artery courses along the central insular sulcus and sends perforating arteries to the sulcus and the adjacent posterior short and anterior long gyri. The stem artery that gives rise to the anterior and posterior parietal arteries courses along the anterior long gyrus (red arrow). An early temporal branch gives rise to the temporooccipital and angular arteries that course on the posterior long gyrus and inferior limiting sulcus. The angular artery turns laterally at the junction of the superior and inferior limbs of the limiting sulci to reach the cortical surface. Ang. = angular; DMCV = deep MCV; M4 = M4 segment of the MCA; Par. = parietal; Perf. = perforating vessel; Pol. = polar; Prefr. = prefrontal; Syl. = sylvian. See legends to Figs. 1, 2, 4, and 5 for definitions of additional abbreviations.
The MCA Trunks and Early Branches
The M2 branches supplying the insula arose from the superior, inferior, and middle trunks of the MCA, the early MCA branches, and an accessory MCA (Fig. 7A and Table 3). The superior or inferior trunk and accompanying branches exclusively supplied different parts of the insula.
Superior Trunk. The stem arteries and cortical branches arising from the superior trunk provided the sole supply to the accessory, transverse, and three short gyri; the anterior limiting and short sulci; and the insular apex in almost all hemispheres (Fig. 7A and Table 3). The superior and inferior trunks supplied the anterior long gyrus and central insular sulcus in similar percentages.
Inferior Trunk. The inferior trunk and its branches supplied the posterior long gyrus, the inferior limiting sulcus, and the limen area in the majority of hemispheres (Fig. 7A). The inferior trunk branches supplied the inferior limiting sulcus in approximately 90% of the hemispheres; however, the early branches contributed to this sulcus in more than half of the hemispheres. The inferior trunk supplied the limen insulae in almost all hemispheres, with minimal contributions from the superior trunk and the early branches.
Early Branches. The early MCA branches—those originating proximal to the bifurcation or trifurcation—have been described in a previous report.27 The early branches, present in 39 of 43 hemispheres, could supply any part of the insula, except the central insular sulcus. The insular areas commonly supplied in part by the early branches included the inferior limiting sulcus, limen area, and anterior limiting sulcus (Fig. 7A and Table 3).
Accessory MCA. The most frequent anomaly of the MCA, the accessory MCA, arises from the ACA. Two accessory MCAs were identified in this study; one supplied the accessory and transverse insular gyri and one supplied the anterior limiting sulcus, prior to terminating in the orbitofrontal area.
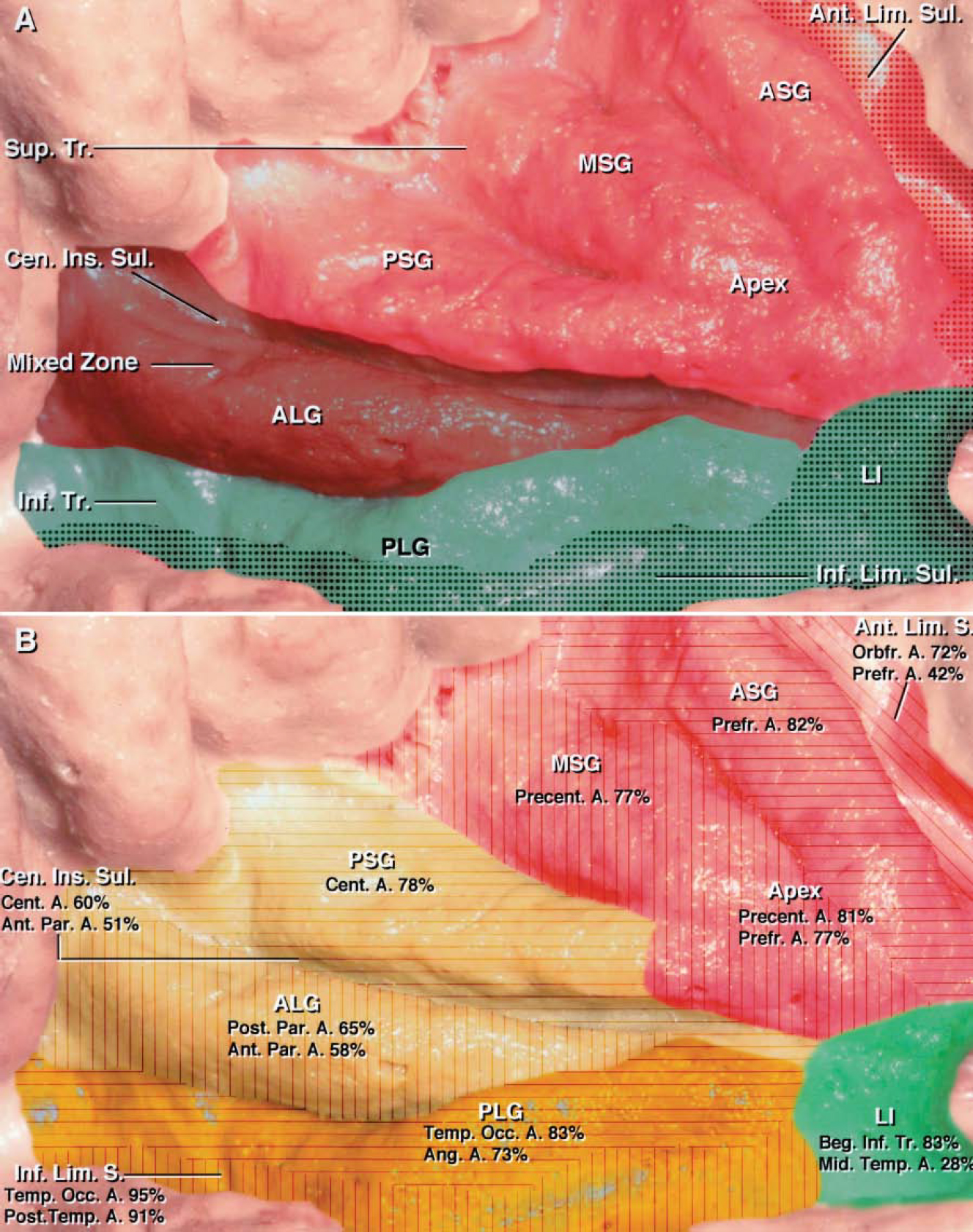
FIG. 7. A: Photograph with superimposed shading to show the arterial supply to the insula (see Table 3). Different portions of the insula were exclusively supplied by the superior or inferior trunk of the MCA, with little overlap. The territory supplied exclusively by the superior trunk of the MCA is shaded red, and the territory supplied exclusively by the inferior trunk is shaded green. Areas receiving a contribution from the early branches are stippled. More specifically, the anterior, middle, and posterior short gyri; the anterior limiting sulcus; the short sulci; and the insular apex were exclusively supplied by the superior trunk of the MCA (red area), with variable contribution from the early branches to the anterior limiting sulcus (stippled area). In contrast to other portions of the insula, the central insular sulcus and anterior long gyrus were supplied by either the superior or the inferior trunk with approximately equal frequency and are referred to as the mixed zone (brown area). The inferior trunk of the MCA supplied the posterior long gyrus, inferior limiting sulcus, and limen area (green area). The inferior limiting sulcus received branches from the inferior trunk in approximately 90% of the cases and from the early branches (stippled area) in more than half of the hemispheres. The limen area received a contribution from the inferior trunk in approximately 90% of the hemispheres and from the early branches in approximately half of the cases (stippled area). B: Diagram of the most common cortical artery contributions to the supply of the insular gyri and sulci. Almost all of the 12 cortical arteries arising from the MCA, with the exception of the temporopolar artery, contributed to the insular supply (see Table 4). Some cortical branches of the MCA arose from a common stem artery that produced two or more cortical arteries. The insular area supplied by the orbitofrontal, prefrontal, and precentral arteries is shaded red; the area supplied by the central and anterior and posterior parietal arteries is shaded yellow; and the area supplied by the angular, temporooccipital, and posterior temporal arteries is shaded orange. The percentages of gyri and sulci in 43 hemispheres supplied by each cortical branch are shown for each specific insular area. The anterior limiting sulcus (oblique lines in red area) received its supply from the orbitofrontal artery in most of the hemispheres. The most consistent areas of supply included the anterior (transverse lines in red area) and middle (vertical lines in red area) short gyri, which were supplied by the prefrontal and precentral arteries in the majority of the hemispheres, respectively. The posterior short gyrus (transverse lines in yellow area) was supplied most frequently by the central artery. The central insular sulcus received perforating arteries from the central artery and the anterior parietal artery in almost half of the hemispheres. The anterior long gyrus (vertical lines in yellow area) was most commonly supplied by the anterior and posterior parietal arteries. The posterior long gyrus (transverse lines in orange area) and the inferior limiting sulcus (vertical lines in orange area) were exclusively supplied by three arteries: temporooccipital, angular, and posterior temporal arteries. The limen insulae (green area) was supplied predominantly by the initial portion of the inferior trunk proximal to the origin of the first cortical artery. The middle temporal artery, arising from an early temporal branch, sent more perforating arteries to the limen insulae than any other cortical artery and supplied it in 30% of the hemispheres. Beg. = beginning; LI = limen insulae; S. or Sul. = sulcus. See legends to Figs. 1 and 5 for definitions of additional abbreviations.

Table 3. Arterial supply of insular areas in 43 cadaveric hemispheres.
Cortical Branches
The 12 cortical arteries arising from the MCA, with the exception of the temporopolar artery, sent branches to the insula (Figs. 5, 6, and 7B and Table 4). These 12 cortical arteries, defined in our previous studies6,27,34 and conforming to definitions proposed by Michotey, et al.,11 are as follows: orbitofrontal, prefrontal, precentral, central, anterior parietal, posterior parietal, angular, temporooccipital, posterior temporal, middle temporal, anterior temporal, and temporopolar arteries. A mean of five cortical arteries (range two–seven arteries) arose from the superior trunk, and four (range two–eight arteries) from the inferior trunk. In the five hemispheres in which the M1 segment ended at a trifurcation, the middle trunks gave rise to either one or two cortical arteries.
The superior trunk and its stem arteries gave rise to most of its cortical branches near the insular pole, within 1 to 2 cm of the MCA bifurcation (Fig. 6). The stem and cortical branches arising from the superior trunks that passed to the anterior cortical areas arose more proximally and had a shorter course along the insula than those arising from the inferior trunk. The first cortical branch of the superior trunk, some of which developed as a stem artery, arose a mean 8.5 mm (range 2.47–17.04 mm) distal to the MCA bifurcation at the insular pole and proximal to the level of the insular apex in most hemispheres. The most common first cortical artery arising from the superior trunk was the orbitofrontal artery, followed in descending order by the prefrontal, precentral, central, anterior parietal, posterior parietal, angular, and temporooccipital arteries.
The stem and cortical branches arising from the inferior trunk ran along the long insular gyri and the inferior limiting sulcus and supplied the posterior part of the insula. The first cortical branch from the inferior trunk arose farther from the MCA bifurcation than those from the superior trunks (Fig. 6). The mean distance between the MCA bifurcation and the origin of the first cortical branch from the inferior trunk was 16 mm (range 3.2–59.63 mm). The most common first cortical branches arising from the inferior trunk included the middle and posterior temporal arteries, followed by the anterior temporal, posterior parietal, and temporopolar arteries. The anterior parietal, central, temporooccipital, and angular arteries infrequently arose as the first branch of the inferior trunk.

Table 4. Cortical arterial supply of insular areas in 43 cadaveric hemispheres.
Cortical Artery Supply of Insular Areas
The orbitofrontal artery supplied the anterior limiting sulcus and the accessory and transverse gyri in the majority of the hemispheres. The prefrontal artery coursed along and supplied the anterior short insular gyrus en route to the junction of the anterior and superior limbs of the limiting insular sulcus in most hemispheres (Fig. 6). Furthermore, it eventually crossed laterally, near the anterior horizontal ramus, to supply the superior aspect of the partes orbitalis and triangularis and most of the middle frontal gyrus. The precentral artery sent branches to the middle short gyrus in the majority of hemispheres. Both the prefrontal and precentral arteries commonly arose from the initial centimeter of the superior trunk (as a common stem) and supplied the region of the insular apex in approximately 80% of the hemispheres (Fig. 7B and Table 4).
The central artery supplied the central insular sulcus in 26 of 43 hemispheres and the posterior short gyrus in 35 hemispheres (Fig. 6). The anterior parietal artery generated perforating arteries along the central insular sulcus in almost half of the hemispheres and contributed to the supply of the posterior short and anterior long gyri. The posterior parietal artery tended to arise from a stem in common with the anterior parietal and central arteries and supplied the anterior long gyrus in 28 hemispheres.
The temporooccipital, angular, and posterior temporal arteries supplied the posterior long gyrus in the majority of hemispheres. The angular artery coursed posteriorly, reaching and sending perforating arteries to the junction of the superior and inferior limiting sulci in most of the hemispheres. The temporooccipital artery arose from a stem in common with the angular artery in the majority of hemispheres, and extended along the inferior limiting sulcus in almost all hemispheres (Fig. 6). The middle temporal artery supplied the limen area in 30% of the hemispheres, sending more branches to the limen insulae than any other cortical artery arising from an inferior trunk or an early temporal branch. The anterior temporal artery supplied only the anterior portion of the inferior limiting sulcus in approximately half of the hemispheres and the limen area in eight hemispheres.
Supply of Insular Sulci and Gyri
Each insular sulcus and gyrus had a distinct pattern of supply from the branches of the MCA (Fig. 7A and B and Tables 3 and 4).
Accessory and Transverse Gyri
In most cerebral hemispheres the stem arteries and the cortical branches arising solely from the superior trunk supplied the accessory and transverse gyri. The cortical arteries arising from the early branches contributed to the supply in one fourth of the hemispheres. The orbitofrontal artery exclusively supplied both gyri, except for a lesser contribution from the prefrontal artery.
Anterior Short Gyrus
Branches arising from the superior trunk supplied the anterior short gyrus. The prefrontal artery most commonly supplied this gyrus.
Middle Short Gyrus
The branches arising from the superior trunk supplied the middle short gyrus in more than 90% of the hemispheres. The precentral artery most often supplied the middle short gyrus, followed by the prefrontal artery.
Insular Apex
Cortical arteries arising from the superior trunk supplied the insular apex in almost all hemispheres. Prefrontal and precentral arteries in similar percentages supplied the apex.
Posterior Short Gyrus
The branches of the superior trunk supplied the posterior short gyrus in most hemispheres. The central artery, followed by the precentral and anterior parietal arteries, most commonly supplied the gyrus.
Central Insular Sulcus and Anterior Long Gyrus
The central insular sulcus and the anterior long gyrus were the only insular areas that received branches from the superior or inferior trunks in similar percentages and are referred to as the “alternated zone” or the “mixed zone” (Fig. 7A and Table 3). There was no overlap in the part of the central insular sulcus and the anterior long gyrus supplied by the superior or inferior trunks. The central and anterior parietal arteries supplied the central insular sulcus with approximately equal frequency. The anterior and posterior parietal arteries supplied the anterior long gyrus in the majority of the hemispheres.
Posterior Long Gyrus
Branches arising from the inferior trunk supplied the posterior long gyrus in 80% of the hemispheres. Angular and temporooccipital arteries exclusively supplied the posterior long gyrus.
Anterior Limiting Sulcus
In approximately 80% of the hemispheres, branches arising from the superior trunk supplied the anterior limiting sulcus. The orbitofrontal and prefrontal arteries exclusively supplied the anterior limiting sulcus.
Inferior Limiting Sulcus
The inferior limiting sulcus received branches arising from the inferior trunk in 90% of the hemispheres and from the early branches in more than half of the hemispheres. The cortical arteries most often supplying the inferior limiting sulcus included the temporooccipital and posterior temporal arteries. The inferior limiting sulcus was rich in perforating branches, having the second highest number of perforating arteries after the central insular sulcus. The perforating arteries were located predominantly along the posterior half of the inferior limiting sulcus.
Limen Area
The limen area was supplied predominantly by the initial portion of the inferior trunk, proximal to the origin of the first cortical artery, in more than 80% of the hemispheres and received a contribution from the early branches in approximately one third of the cases. The middle temporal artery supplied the limen area in 30% of the hemispheres, sending more perforating branches to the limen insulae than any other cortical artery arising from the inferior trunk or an early branch.
Large Perforating Arteries of the Insula
Tiny perforating arteries arose from all of the MCA trunks and branches as they crossed the insula. In this study we focused on the larger perforating branches. A total of 194 insular perforating branches equal to or larger than 0.3 mm in diameter (mean 4.5 branches per hemisphere) arose from the MCA trunks, the early branches, and the cortical and stem arteries as they crossed the insula. The only cortical branch that did not send perforating branches to the insula was the temporopolar artery. The branches larger than 0.3 mm in diameter most commonly arose from the central, angular, and posterior parietal arteries and penetrated the posterior half of the central insular and inferior limiting sulci and the long gyri (Fig. 6 and Tables 5 and 6). Twenty percent of the 194 larger insular perforating arteries had a diameter greater than 0.5 mm and were directed predominantly to the posterosuperior part of the long gyri.

Table 5. Distribution of 194 larger perforating arteries of the insular cortex.

Table 6. Origin of 194 larger insular perforating arteries.
Venous Relationships
Analysis of data obtained in this study revealed numerous exceptions to the traditional concept that the superficial venous system of the cerebrum drains the superficial cortical areas bordering the sylvian fissure and that the deep venous system drains the insula.15,19,38 In addition, numerous anastomoses between the two systems were demonstrated.
Superficial Sylvian Vein
The SSV is the largest vein draining along the posterior ramus of the sylvian fissure, and the deep MCV is the predominant vein draining the insula. The SSV arose at the posterior end of the sylvian fissure in 38 of 43 hemispheres, was absent in two, and was hypoplastic posterior to the central sulcus in three (Fig. 8). It usually arose as a single trunk (37 hemispheres), but might also arise as two trunks that join before emptying into the venous sinuses along the sphenoid ridge. The SSV coursed downward and forward, usually on the temporal side of the posterior ramus of the sylvian fissure (31 hemispheres). It turned medially at the anterior end of the sylvian fissure and emptied into the part of the sphenoparietal sinus coursing just below the medial part of the sphenoid ridge in 35 hemispheres. In the remaining hemispheres it emptied directly into the cavernous sinus or into a sphenopetrosal sinus, which crossed the floor of the middle fossa to empty into the superior petrosal sinus. The SSV and deep MCV emptied together into the same point at the junction with the sphenoparietal sinus in nine hemispheres and emptied separately into the sinus in 11 hemispheres. The mean distance between the limen insulae, and the junction of the SSV and the sphenoparietal sinus was 24.8 mm (Table 1).
The veins draining the frontal, parietal, and temporal lobes along the posterior ramus of the sylvian fissure are called “frontosylvian,” “parietosylvian,” and “temporosylvian” veins, respectively. The SSV received a mean of six frontosylvian, four parietosylvian, and five temporosylvian veins along its course on the posterior ramus of the sylvian fissure (Fig. 8). The frontosylvian veins drained the inferior frontal gyrus, adjoining a portion of the middle frontal gyrus and the inferior portion of the precentral gyrus. The frontosylvian veins descended on the frontal operculum, passing anteriorly or anteroinferiorly in all but a few hemispheres to reach the SSV. In the hemispheres in which the frontosylvian veins did not drain into the SSV, vessels that emptied through the vein of Trolard into the superior sagittal sinus drained the area. The frontosylvian veins had more frequent anastomotic connections with the insular veins and drained more of the insula than either the parietosylvian or temporosylvian veins. Furthermore, the frontosylvian veins provided the venous drainage for the middle short gyrus, and the insular apex in 80% of the hemispheres. Other areas drained in part by the frontosylvian veins included the anterior and posterior short gyri; the anterior long gyrus; and the central, precentral, and anterior limiting sulci.
The parietosylvian veins drained the postcentral gyrus and inferior parietal lobule and either descended to join the SSV or ascended to join the veins entering the superior sagittal sinus with approximately equal frequency (Fig. 8). Occasionally, the parietosylvian veins emptied into the upper end of the vein of Labbé or into the vein of Trolard. The parietosylvian veins contributed to venous drainage of the anterior long gyrus and central insular sulcus in two hemispheres.
The areas drained by the temporosylvian veins were larger than those drained by the fronto- and parietosylvian veins (Fig. 8). The temporosylvian veins extended along the superior temporal gyrus, from the temporal pole to the posterior end of the sylvian fissure. They joined the SSV in 15 hemispheres and both the SSV and the vein of Labbé in the remaining hemispheres. Furthermore, the temporosylvian veins contributed to venous drainage of the posterior long gyrus and inferior limiting sulcus in some hemispheres (Tables 7 and 8).

FIG. 8. Photographs obtained during stepwise dissections of the right cerebral hemispheres, showing the venous drainage of the opercular and insular areas (A–D). A: Lateral view. The portion of the SSV posterior to the central sulcus is small, and the anterior portion is large. The SSV receives drainage from the frontosylvian and temporosylvian veins and empties into the sphenoparietal sinus along the sphenoid ridge. The frontosylvian veins drain the inferior frontal gyrus and descend along the anterior horizontal and anterior ascending rami of the sylvian fissure to join the SSV. The temporosylvian veins extend along the superior temporal gyrus. The more posterior temporosylvian veins join into the vein of Labbé. B: Enlarged lateral view of the sylvian fissure. The frontosylvian veins drain the inferior frontal gyrus and descend along the anterior horizontal and anterior ascending rami to join the SSV. The temporosylvian veins cross the superior temporal gyrus to join the SSV. C: Lateral view. The sylvian fissure has been opened to expose the relationship between the SSV and the insular drainage. Frontosylvian tributaries of the SSV drain a portion of the middle and posterior short insular gyri. The frontoorbital ramus, the least frequently occurring of the four rami of the sylvian fissure, is located inferior to the anterior horizontal ramus and is crossed by the frontoorbital vein. D: Enlarged lateral view of Fig. 8C. Drainage of the insular apex and middle short gyrus occurs predominantly through the frontosylvian tributaries of the SSV. Photographs obtained during stepwise dissections of right cerebral hemisphere to display the relationship between the SSV and insular veins (E and F). E: Lateral view. The frontosylvian and parietosylvian veins empty into the SSV. The temporosylvian veins cross the superior temporal gyrus to empty predominantly into the vein of Labbé. F: The frontal and temporal opercula have been retracted to expose the connections between the SSV and insular veins. The precentral insular vein drains the posterior short gyrus and empties into the SSV. The frontosylvian tributaries of the SSV drain the middle short gyrus and insular apex. The anterior insular vein drains the anterior short gyrus and descends to terminate into the deep MCV near the limen area. Trib. = tributaries. See legends to Figs. 1, 2, 5, and 6 for additional abbreviations.

Table 7. Patterns of superficial (SSV) and deep (deep MCV) venous drainage of insular areas.
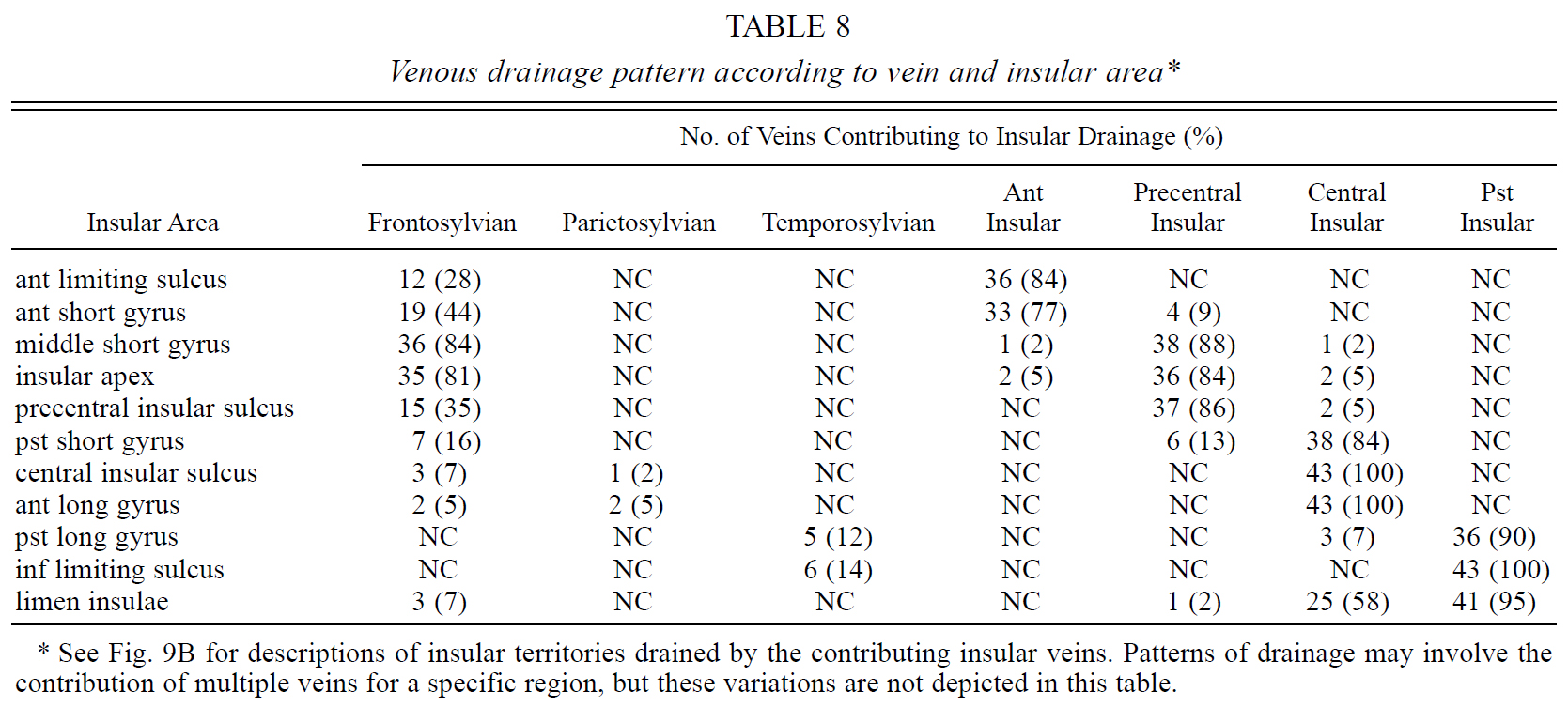
Table 8. Venous drainage pattern according to vein and insular area.
Insular Veins
The veins draining the insula emptied predominantly into the deep MCV in almost all hemispheres; however, the tributaries of the SSV drained some insular areas. There were also anastomoses between the insular veins and the superficial venous system in most hemispheres (Fig. 9A and Table 7).
Insular drainage was categorized into three groups based on whether the area drained into the superficial, deep, or both venous systems: superficial, deep, and transitional (Fig. 9A and Table 7). The limen area, inferior limiting sulcus, long gyri, and central insular sulcus drained predominantly into the deep venous system. The middle short gyrus and insular apex drained predominantly into the SSV. The transitional zone, which could be drained by both venous systems, included the anterior and posterior short gyri and the anterior limiting sulcus. These transitional areas were more frequently drained by the deep venous system than the superficial one (Fig. 9A).
The insular veins have been named according to their relationship with the insular sulci and gyri.7,25,36,37 Four insular veins were identified in our study: anterior, precentral, central, and posterior veins (Figs. 9B and 10). These veins demonstrated distinct characteristics in regard to their origin, course, relationship to the SSV, area of drainage, and termination patterns.

FIG. 9. A: Diagram of the insular areas drained by the superficial and deep venous systems in 43 hemispheres. See Table 7 for a listing of the insular areas drained by the superficial and/or deep venous systems. The territory drained by the deep venous system is shaded purple, whereas that drained by the superficial venous system is light blue. The limen area, inferior limiting sulcus, long gyri, and central insular sulcus are drained by the deep MCV (purple area). The SSV drained the middle short gyrus and insular apex more commonly than any other insular area (light blue area). The transitional zone, which is usually drained by both the superficial and deep venous systems, included the anterior and posterior short gyri and the anterior limiting sulcus (stippled area). These transitional areas emptied three to eight times more frequently into the deep MCV than the SSV. B: Diagram of areas drained by insular veins. See Table 8 for a listing of the insular areas drained by the four insular veins. The territory drained by each insular vein is shaded a different color: anterior insular vein (dark green), precentral insular vein (light green), central insular vein (light blue), and posterior insular vein (dark blue). The areas having significant anastomotic connections with the SSV are stippled. The anterior insular vein drained the anterior limiting sulcus and the anterior short gyrus (dark green area). Although the anterior short gyrus was drained by the anterior insular vein in the majority of hemispheres, the frontosylvian veins contributed to drainage in almost half of the cases (stippled area). The precentral insular vein drained the middle short gyrus and insular apex and displayed more anastomoses with the SSV than any other insular vein. The frontosylvian veins contributed to drainage of the middle short gyrus and insular apex in more than 80% of the hemispheres (stippled area). The central insular vein drained the posterior short and anterior long gyri and the central insular sulcus in almost 90% of the specimens (light blue). The posterior insular vein exclusively drained the posterior long gyrus and the inferior limiting sulcus (dark blue). The most complicated pattern of insular drainage was in the limen area (light and dark blue areas), which was drained by a combination of the central and posterior insular veins in more than half of the cases and by the posterior insular vein alone in the other 40% of cases. The limen area is the only one drained almost solely by the deep venous system without any contribution from the SSV. See legends to Figs. 1, 5, and 7 for additional abbreviations.
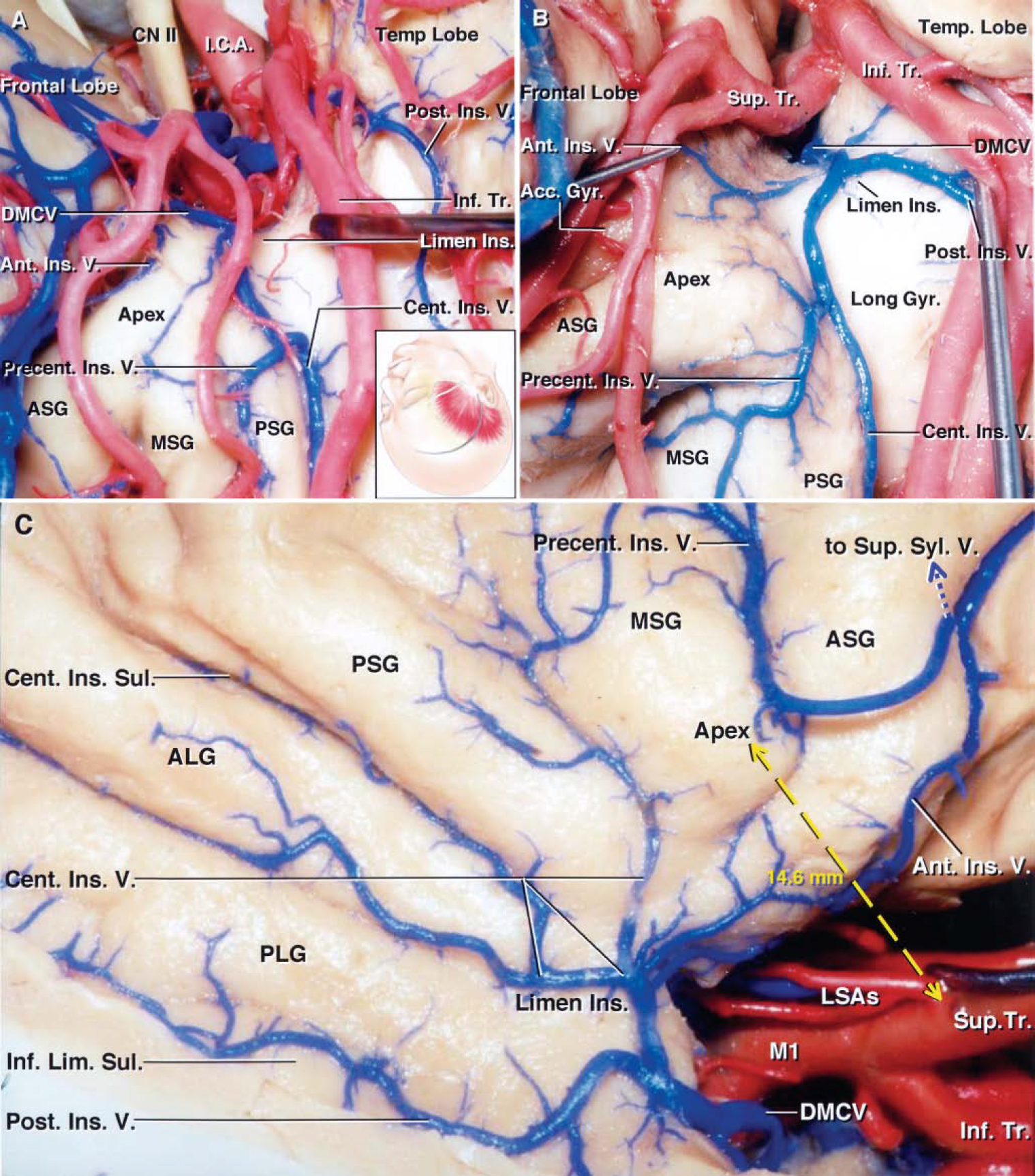
FIG. 10. Photographs obtained during cadaveric dissection. A: Right pterional exposure. The inset shows the position of the head and skin incision for the frontotemporal craniotomy. The sylvian fissure has been widely opened and the frontal and temporal opercula have been retracted to expose the anterior half of the insula. The inferior trunk has been retracted to expose the junction of the central and precentral insular veins forming a common stem. The central insular vein courses along the central insular sulcus and drains the sulcus and the adjacent portion of the anterior short gyrus. The precentral insular vein drains the middle and posterior short gyri. The posterior insular vein courses along the inferior limiting sulcus and joins the deep MCV in the limen area. B: Another right pterional exposure. The M2 branches have been retracted to expose the formation of the deep MCV. The precentral insular vein drains the middle short gyrus and insular apex and joins the central insular vein behind the apex. The central insular vein courses along the central insular sulcus and joins the precentral insular vein to form a common trunk, which receives the anterior and posterior insular veins forming the deep MCV at the limen insulae. C: Lateral view of right cerebral hemisphere. The frontoparietal and temporal opercula have been retracted to expose the course of the insular veins. The four insular veins and the initial portion of the deep MCV have been exposed. The anterior insular vein courses downward and backward near the anterior limiting sulcus, drains the anterior limiting sulcus and anterior short gyrus, and empties into the deep MCV. The precentral insular vein, the insular vein most commonly connected with the SSV, drains the middle short gyrus and the insular apex, crosses the anterior short gyrus, and turns superficially to empty into the SSV without contributing to the formation of the deep MCV. The central insular vein courses anteroinferiorly; drains the central insular sulcus, the posterior short and anterior long gyri, and the limen area; and joins the posterior insular vein. The posterior insular vein courses anteriorly along the inferior limiting sulcus; drains the inferior limiting sulcus, the adjacent portion of the posterior long gyrus, and the limen area; and joins the central insular vein to form the initial portion of the deep MCV near the limen insulae. The bifurcation of the MCA has been retracted to expose the LSAs arising from the postbifurcation trunks. Although the most lateral LSA most frequently originates from the prebifurcation M1 segment, in this particular hemisphere the artery originates from the superior trunk. The mean distance from the insular apex, the most laterally prominent or highest point on the insula, to the origin of the most lateral LSA was less than 15 mm (yellow dashed arrow). See legends to Figs. 1, 4, 5, and 6 for definitions of abbreviations.
Anterior Insular Vein
The anterior insular vein was found in 36 of 43 hemispheres. It proceeded downward and backward, on or near the anterior limiting sulcus, and terminated in the deep MCV near the limen insulae in 34 hemispheres and into the SSV in two hemispheres (Fig. 10). This vein emptied directly into the deep MCV, independent of the other insular veins, in almost half of the hemispheres. It drained the anterior limiting sulcus and the anterior short gyrus in the majority of hemispheres. In addition, it had anastomoses with the tributaries of the SSV, most frequently the frontosylvian veins.
Precentral Insular Vein
The precentral insular vein, found in 39 of 43 hemispheres, coursed straight anteroinferiorly, usually on the precentral insular sulcus, and drained the middle short gyrus and insular apex in most hemispheres (Fig. 10). It joined the central and posterior insular veins and terminated in the deep MCV in 32 hemispheres. In the remaining hemispheres, it coursed superficially to join the SSV (Figs. 8 and 10). It had anastomoses with the tributaries of the SSV in approximately half of the hemispheres, most commonly the frontosylvian tributaries. Areas drained by the precentral insular vein were in fact drained by the tributaries of the SSV more often than any other insular area (Fig. 9B and Table 8).
Central Insular Vein. The central insular vein, present in all 43 hemispheres, coursed anteroinferiorly along the central insular sulcus (Fig. 10). It provided the sole means of drainage for the posterior short and anterior long gyri and the central insular sulcus in almost 90% of the hemispheres. The central insular vein together with the posterior insular vein contributed to the drainage of the limen area in almost half of the hemispheres. The central insular vein emptied into the deep MCV in 42 of 43 hemispheres; in one hemisphere, the vein joined the SSV along the posterior ramus of the sylvian fissure. The frontosylvian or parietosylvian veins had anastomoses with the central insular vein and contributed to the drainage of insular areas normally drained by the central insular vein in a few hemispheres.
Posterior Insular Vein. The posterior insular vein, present in all 43 hemispheres and referred to as the “first portion of the deep MCV,”35 most frequently joined with the central insular vein to form a common stem that ran medially to form the deep MCV in almost all of the hemispheres (Fig. 10). It coursed along the long gyri, usually near or along the inferior limiting sulcus, and drained the inferior limiting sulcus, posterior long gyrus, and limen area in almost all hemispheres (Figs. 9B and 10 and Table 8). Occasionally, it had anastomoses with the temporosylvian veins.
Patterns of Drainage of the Insular Areas
Each insular area had a distinct drainage pattern (Fig. 9B and Table 8).
Anterior Short Gyrus
In nearly 80% of the hemispheres, the anterior insular vein alone or in part drained the anterior short gyrus. The frontosylvian veins contributed to the drainage of this gyrus in almost half of the hemispheres (Fig. 9B).
Middle Short Gyrus and Insular Apex
The middle short gyrus and insular apex were drained predominantly by the precentral insular vein, with the frontosylvian tributaries of the SSV contributing to drainage in most of these hemispheres.
Posterior Short Gyrus
In the majority of hemispheres, the central insular vein drained the posterior short gyrus; in six hemispheres, the precentral insular vein drained this gyrus.
Central Insular Sulcus and Anterior Long Gyrus
The central insular vein exclusively drained the central insular sulcus and the anterior long gyrus.
Posterior Long Gyrus
In most hemispheres the posterior insular vein drained the posterior long gyrus; the central insular vein drained this gyrus in only three hemispheres.
Anterior Limiting Sulcus
In the majority of hemispheres (80%), the anterior insular vein drained the anterior limiting sulcus. The frontosylvian veins contributed to sulcal drainage in 12 hemispheres.
Inferior Limiting Sulcus
The inferior limiting sulcus was drained by the posterior insular vein in all hemispheres, with a contribution from the temporosylvian veins in six hemispheres.
Limen Area
The limen area was drained by the combination of the central and posterior insular veins in almost half of the hemispheres and by the posterior insular vein alone in the remaining hemispheres.
Deep MCV
The deep MCV is the largest and most consistent tributary of the basal vein.7,15,24,36 It passes medially across the anterior perforated substance where it is joined by the anterior cerebral vein to form the anterior segment of the basal vein, which ends at the anterolateral portion of the cerebral peduncle where it receives the peduncular vein (Fig. 11). The anterior segment of the basal vein drains the basal areas anterior to the brainstem, including the optic chiasm, the anterior perforated substance, the orbital surface of the frontal lobe, and the insula.
The deep MCV was formed by the union of insular veins near the limen insulae in all hemispheres. Usually, it is described as being formed by the union of the anterior, precentral, central, and posterior insular veins at the limen area to form a common transverse trunk.15,36 This pattern was found in only 27% of the hemispheres, however. The anterior insular vein joined the deep MCV independently, distal to the union of the central and posterior insular veins, in 42% of the hemispheres. One of the four insular veins, most commonly the precentral insular vein, may infrequently empty into the SSV and not the deep MCV (Fig. 10).
We observed several variations in the course and termination of the deep MCV (Figs. 11 and 12). In 23 hemispheres, the deep MCV crossed the limen insulae and passed between the proximal M1 segment and the anterior perforated substance to join other veins in forming the initial portion of the basal vein (Fig. 11). The deep MCV was usually joined by the frontoorbital vein prior to reaching the anterior perforated substance. As it crossed the anterior perforated substance it was joined by the olfactory and inferior striate veins. The anterior cerebral vein subsequently joined the deep MCV to form the anterior segment of the basal vein in 23 hemispheres; in the remaining 20 hemispheres, the deep MCV and some of the tributaries of the initial segment of the basal vein formed a common stem, which coursed forward to empty into the sphenoparietal sinus (Figs. 11 and 12). The common stem, most often formed by the combination of the deep MCV with the olfactory and frontoorbital veins, ran parallel to the SSV and sometimes received the inferior striate and anterior cerebral veins prior to entering the sphenoparietal sinus (Fig. 12).

FIG. 11. Photographs obtained during cadaveric dissection, showing variations in the termination pattern of the deep MCV. A: Anterosuperior view. The anterior portions of both cerebral hemispheres have been removed to expose the temporal horns, basal cisterns, deep MCVs, basal veins, and PCAs. The PCAs arise from the basilar artery in the interpeduncular cistern and pass around the brainstem. The anterior choroidal arteries course around the left and right unci to reach the temporal horns. The right deep MCV passes around the limen area to join the basal vein. The right basal vein passes above the PCA in the crural and ambient cisterns. The anterior segment of the basal vein ends where the peduncular vein joins the basal vein at the anterolateral portion of the cerebral peduncle. The inferior choroidal vein joins the basal vein at the posterior edge of the cerebral peduncle. On the left side, the anterior segment of the basal vein is hypoplastic and the deep MCV courses below the M1 segment and empties into the sphenoparietal sinus along the sphenoid ridge. The middle segment of the basal vein passes around the brainstem above the PCA and is joined by the inferior ventricular and choroidal veins. Both basal veins join the vein of Galen in the quadrigeminal cistern. Views of the deep MCV draining into a left basal vein (B and C). B: Lateral view of the anteroinferior portion of the left insula. The frontoparietal and temporal opercula have been retracted. The MCA bifurcation has been retracted anteriorly to expose the insular veins. The precentral insular vein joins the SSV. The central and posterior insular veins join at the limen insulae to form the deep MCV. The anterior insular vein courses near the anterior limiting sulcus and joins the deep MCV. C: Anteroinferior view of the anterior perforated substance of the left hemisphere shown in B. The prebifurcation M1 segment and the LSAs have been retracted to expose the formation of the anterior segment of the basal vein. The deep MCV passes medially across the limen insulae and anterior perforated substance and is joined by the inferior striate and olfactory veins. The anterior segment of the basal vein begins at the union of the deep MCV and the anterior cerebral vein below the anterior perforated substance. D: Inferior view of the left frontal lobe and anterior perforated substance in another hemisphere. The deep MCV is formed by the union of the insular veins and receives the inferior striate veins as it crosses the anterior perforated substance. The basal vein begins at the junction of the anterior cerebral vein, deep MCV, and olfactory vein. Parahip. = parahippocampal; Ped. = peduncular; Vent. = ventricular. See legends to Figs. 1 and 4 to 6 for definitions of additional abbreviations.

FIG. 12. Photographs obtained during cadaveric dissection, showing the deep MCV and SSV at their junction with the sphenoparietal sinus. Right pterional exposure (A and B). A: The inset shows the position of the head and skin incision. The sylvian fissure has been opened to expose the anteroinferior portion of the insula, the MCA bifurcation, and the M2 branches distal to the limen insulae. The deep MCV courses downward and forward and receives the frontoorbital vein prior to emptying into the sphenoparietal sinus. The SSV passes superior to the deep MCV as a separate trunk that empties into the sphenoparietal sinus with the deep MCV. B: The MCA bifurcation has been retracted to expose the central and posterior insular veins as they join to form the deep MCV, which courses toward the sphenoid ridge to empty into the sphenoparietal sinus. Right pterional exposure in another specimen (C and D). C: The sylvian fissure has been opened to expose the anterior half of the insula. The MCA bifurcation is located at the level of the limen insulae. An early temporal branch arises from the prebifurcation M1 segment. The postbifurcation trunks have been retracted to expose the central and posterior insular veins which cross the limen insulae and join to form the deep MCV, which empties into the sphenoparietal sinus. The SSV also empties into the sphenoparietal sinus. The frontosylvian tributaries of the SSV drain the short gyri and insular apex. D: The SSV has been retracted to the temporal side to show the relationship between the frontoorbital vein and deep MCV. The frontoorbital vein descends from the posterior orbital gyrus of the frontal lobe, crosses the early temporal branch of the MCA, and joins a deep MCV that empties into the sphenoparietal sinus. Sph. = spheno or sphenoid. See legends to Figs. 1, 2, 4 to 6, and 8 for definitions of other abbreviations.
Discussion
The frequency with which the sylvian fissure is opened, thus exposing the insula, increases the importance of understanding the anatomical features of the insula and its relationship to the sylvian fissure. Despite the frequency with which neurosurgeons encounter the insula, few authors have described the anatomy of the insular region and its arterial and venous components.2,10,20,28,29,32
Insular Exposure
The most common insular exposure is directed to the anteroinferior portion near the pole and limen insulae for exposure of the ICA, MCA bifurcation, circle of Willis, basal cisterns, medial sphenoid ridge, and adjacent skull base. Exposing the limen medially along the basal cisterns and skull base requires that the approach be directed downward from the area below the pars triangularis along the medial side of the temporal operculum to the insula. The MCA bifurcation and origin of the trunks are located medial to the temporal operculum, but numerous MCA branches distal to the bifurcation, especially those from the superior trunk, are located in the upper portion of the insular pole and medial to the frontoparietal operculum. Lesions involving the insular cortex and the underlying white and gray matter, such as tumors or AVMs, require a wider exposure of the insula together with opening of the insular compartments medial to both the frontoparietal and temporal opercula.
To expose the insular region fully, the sylvian fissure must be widely split.9,42 The area below the apex of the pars triangularis, where the three major rami of the sylvian fissure converge, provides the widest corridor to open the anterior end of the sylvian fissure. The distance along the cortical surface from the apex of the pars triangularis to the superior limiting sulcus of the insula is less than 2 cm. In 20% of the hemispheres, the pars triangularis is small and the tip retracted upward away from the fissure so that the anterior horizontal and anterior ascending rami arise as a common stem, providing an extra 10 mm of width below the apex of the pars triangularis to begin the opening into the sylvian fissure (Fig. 1).
Different parts of the insula can be approached more or less directly by preferentially opening specific portions of the sylvian fissure. Opening to the depths of the upper end of the anterior ascending ramus of the sylvian fissure exposes the superior limiting sulcus and the adjacent portion of the anterior or middle short gyri in the majority of the hemispheres (Fig. 2). Opening to the depth of the posterior end of the posterior ramus exposes the junction of the superior and inferior limiting sulci at the posterosuperior edge of the insula. The anterior horizontal ramus has a less consistent relationship to the underlying limiting sulci. Opening to the depths of the anterior end of the anterior horizontal ramus may expose the superior limiting sulcus, the anterior limiting sulcus, or, most often, the junction of the two sulci together with the adjacent portion of the upper margin of the anterior short gyrus.
When opened and followed to the depths of its posterior end, the frontoorbital ramus, identified in 63% of the hemispheres, exposes the midportion of the anterior limiting sulcus along with the adjacent part of the anterior short and accessory insular gyri (Fig. 2). The frontoorbital ramus has previously been categorized as a part of the orbital sulci rather than a ramus of the sylvian fissure, even though it opens into the fissure.12,14,28 From a neurosurgical standpoint, we believe that this sulcus should be considered as part of the sylvian fissure,36 because it opens into the fissure as does the anterior ascending and anterior horizontal rami. The frontoorbital vein coursed along the ramus in most hemispheres. The ramus can commonly be opened without sacrificing the vein. Opening the frontoorbital ramus, when present, significantly improved the exposure of the anterior limiting sulcus as well as the accessory and transverse gyri of the insula (Fig. 1). Opening along the frontoorbital ramus may reduce both the amount of retraction of the inferior frontal gyrus needed to reach the anteroinferior portion of the insula and the incidence of motor and speech dysfunction associated with retraction of the inferior frontal gyrus.
The distance from the cortical surface along the opercular lips to the superior and inferior limiting sulci varies from the anterior to posterior portion of the fissure. Anteriorly, the distance to the superior limiting sulcus is approximately 2 cm and is always greater than the distance to the inferior limiting sulcus. At the posterior end of the fissure, the distance to the superior and inferior limiting sulci are nearly identical (mean 2.5 cm).
Relationship Between the Limen Insulae and the Anterior Perforated Substance
Yasargil, et al.,43 asserted that the medial aspect of insular tumors may envelop the LSAs and that the medial extent of resection of these tumors should not extend medial to the LSAs. Lang, et al.,9 also posited that the medial extent of resection of insular tumors should be limited to the area lateral to the most lateral LSA. An awareness of the location of the most lateral LSA may be helpful during surgery for insular tumors, because motor deficits such as hemiparesis due to obliteration of these perforating arteries make up a significant number of the complications that occur following insular tumor surgery.5,9,43,45 The most lateral LSA originated most commonly from the main trunk of the M1 segment, distal to the early branches; the second most common site of origin was from the inferior trunk. The distance from the insular apex to the origin of the most lateral LSA was approximately 15 mm (Fig. 10).
The width of the limen insulae, defined as the distance between the anterior end of the long gyri and the middle of the posterior orbital gyrus, was a mean of 2 cm. Analysis of previous reports has demonstrated that the limen insulae should be considered as the lateral limit of the anterior perforated substance.34 Given the importance of the position of the LSAs during surgery, we believe that the point of entrance of the most lateral LSA should be considered as the lateral limit of the anterior perforated substance. A recess lies between the medial border of the limen insulae and the lateral limit of the anterior perforated substance, which is the point of entrance of the most lateral LSA. We refer to this area as the “limen recess” (Fig. 4). Importantly, the limen recess, which had a mean width of 15.3 mm, was devoid of perforating arteries in all the cerebral hemispheres studied. These findings indicate that injuries to LSAs can be avoided if the medial resection plane of an insular tumor does not extend beyond the limen recess.
Arterial Supply
Cortical MCA Branches
The contribution of the individual cortical branches of the MCA to insular blood supply has not been studied previously. Awareness of which M4 cortical branch originates from which stem artery along the insula and knowledge of the most common arterial vascularization patterns of insular gyri and sulci may be helpful in the surgical planning of approaches along the sylvian fissure and insula.
The cortical MCA branches usually arise from one of five parent vessels: superior, inferior, or middle trunks of the MCA, early branches, or infrequently from an anomalous vessel such as an accessory MCA. Previously, we described the similarities between the MCA anomalies and the early branches and noted that accessory MCAs typically supply the accessory and transverse gyri of the insula and terminate in the orbitofrontal territory.27 The early branches provide a more extensive supply to the insula than the anomalous vessels, however. The early branches may supply any portion of the insula, except for the central insular sulcus.
It has been reported that the superior trunk of the MCA is responsible for the blood supply to the entire insula in 50% of cases.32In the present study, however, we found a more significant role for the inferior trunk and the early branches. The superior trunk provided the predominant supply to the accessory and transverse gyri, the short gyri of the insula, the insular apex, and the anterior limiting sulcus, with only a minimal contribution from the inferior trunk (Fig. 7A). On the other hand, the inferior trunk generally supplied the posterior long gyrus, inferior limiting sulcus, and limen insulae. The central insular sulcus and anterior long gyrus formed a separate middle zone that was supplied by either the superior or inferior trunk, without any contribution from the other trunk. The central insular zone displayed a mixed, but not overlapping, pattern of supply more commonly than other parts of the insula. We concur with previous study data indicating that there is minimal overlap in the insular areas supplied by the superior and inferior trunks. Each of the insular gyri and sulci had a characteristic pattern of supply by the cortical arteries as described in Results (Fig. 7B).
Insular Perforating Arteries
The majority of insular perforating arteries can be coagulated to devascularize intrinsic tumors and vascular malformations without damaging the putamen and the internal capsule.41 Nonetheless, it has been reported that the interruption of blood flow to the larger insular perforating arteries during resection of intrinsic insular tumors may result in hemiparesis and that these arteries should be preserved to prevent infarction of the corona radiata.9,29 Yasargil and colleagues41–43 asserted, and we agree, that the larger insular perforating arteries most commonly arise from the distal portion of the M2 branches, and they recommended avoiding obliteration of these larger-caliber insular arteries. The larger insular perforating arteries were most commonly located on the posterior half of the central insular sulcus and on the long gyri. Varnavas and Grand32 observed a large insular perforating artery at the junction of the superior and inferior limiting sulci in 25% of the uninjected hemispheres in their study. Contrary to previous reports, the larger perforating arteries were identified not only in the posterosuperior portion of the insula, but also posteroinferiorly.
Venous Drainage
Complete angiographic delineation of the drainage of the sylvian fissure and the insula has proven difficult because the SSV and the deep MCV fill more or less simultaneously and drain into multiple sites.26,38 Moreover, the variations in the number, size, course, connections, and drainage patterns of the SSV and deep MCV are considerably more complex than the variations in the arterial supply.13,19 The relationships and communications between the SSV and deep MCV and their contribution to the venous drainage of the insular and opercular areas have not been clearly described in the literature.
The sylvian fissure will usually be easier to split by working above, rather than below, the SSV because the vein courses approximately 4 mm below the fissure in more than 80% of hemispheres. The drainage pattern of the tributaries of the SSV varies considerably because these tributaries of ten have connections to the veins of Trolard and Labbé. The SSV drains into the sphenoparietal sinus in more than 85% of the cases; however, it can also drain directly into the cavernous sinus or the sphenopetrosal sinus.
There have been no previous reports detailing the drainage patterns of the different parts of the insula. The insular veins are usually described as being part of the deep venous drainage system, although it has been recognized that the superficial venous system may participate in insular drainage.14,19,32,37 The insular veins emptied predominantly into the deep MCV in our study, but there was significant drainage to the SSV in most hemispheres. All of the insular veins have connections to the superficial venous system. Contrary to previously published data, drainage of the middle short gyrus and insular apex was predominantly directed laterally into the SSV. Several other areas, including the posterior short gyrus and the anterior limiting sulcus, formed transitional zones that drained into both the superficial and the deep venous systems (Fig. 9A). The precentral insular vein, which usually drains the middle short gyrus and the insular apex, is the one that most commonly empties into the superficial venous system.
Preoperative mapping of the venous drainage of the insula may be important not only for tumoral lesions of the insular region, but also for vascular disorders. Most AVMs in the sylvian fissure are fed by small, short branches of arteries that must be preserved because they continue to supply blood to normal brain tissue distal to the AVM. Zimmerman, et al.,46 reported a series of pure sylvian fissure AVMs and asserted that these AVMs should be resected via an anteroposterior or posteroanterior approach, unlike most parenchymal AVMs, which are removed in circumferential dissection. These authors proposed a resection plane according to whether drainage was directed posteriorly through the cortical veins or anteriorly through the deep MCV into the sphenoparietal sinus. The AVM was removed in a proximal to distal direction along the sylvian fissure if venous drainage was directed posteriorly and in a distal to proximal direction if drainage was directed anteriorly.46 In approximately 45% of the hemispheres, the deep MCV coursed anteriorly to empty into the sphenoparietal sinus; half of these cases had a connection to the SSV. A detailed knowledge of the venous anatomy of the insula should help in planning resections of vascular anomalies along the sylvian fissure.
Embryological formation of the basal vein results from sequential changes involving anastomoses, deletions, and reanastomoses of primitive pial venous plexuses.7,17,36 Therefore, it is not surprising that there are many variations in the formation of the initial portion of the basal vein and deep MCV. We observed that the deep MCV drained into either the sphenoparietal sinus or the basal vein. In nearly half of the hemispheres studied, the deep MCV passes medially and joins other veins to form the initial segment of the basal vein. In the other half, the deep MCV courses forward, accompanying the SSV, to empty into the sphenoparietal sinus (Fig. 11). The fact that a large number of deep MCVs empty into the sphenoparietal sinus, as opposed to the basal vein, could be due to the regression of the longitudinal anastomoses between the first and second segments of the basal vein (Fig. 12). The tributaries of the basal vein that join the deep MCV and those of the common stem that drain into the sphenoparietal sinus may limit retraction of the frontal and temporal lobes during pterional and pretemporal approaches.
Obliteration of the SSV and deep MCV along the sphenoid ridge may cause seizures and a facial palsy plus aphasia if the occlusion occurs on the left side.21 The number of tributaries to be sacrificed should be kept to a minimum because of possible undesirable sequelae. Increasing venous diameter and size of the drainage area increases the likelihood of ill effects if the vein is sacrificed. The central and posterior insular veins have the largest diameter and drainage area of all the insular veins.
Transinsular Approaches to the Temporal Horn and Basal Cisterns
The transinsular route through the pterional–transsylvian approach has been used for selective amygdalohippocampectomy39,44 and adapted for lesions located in the ambient cistern.41 An understanding of the relationship between the insular cortex and the deep landmarks, such as the choroidal fissure, is helpful in planning transinsular approaches (Fig. 13).21
This approach involves opening the sylvian fissure and exposing the anterior one third of the insula down to the inferior limiting sulcus and the initial 2 cm of the M2 segment (Fig. 14). A cortical incision of approximately 1.5 to 2 cm in length in the inferior limiting sulcus of the insula provides access to the temporal horn of the lateral ventricle. The arteries running along the inferior limiting sulcus commonly arise from the early branches of the M1 segment. The anterior one half of the inferior limiting sulcus was found to be devoid of larger perforating arteries in all of the hemispheres studied. To perform the transinsular approach, a cortical incision is made through the inferior limiting sulcus approximately 1 to 2 cm posterior to the point at which the anterior end of the long gyri fuses with the temporal pole. The incision is made lateral to the inferior trunk between the M2 branches, most commonly the anterior and middle temporal arteries, which run parallel to the inferior limiting sulcus. The cortical incision along the inferior limiting sulcus can be located medial or lateral to the posterior insular vein without sacrificing the vessel. The anterior and middle portions of the temporal horn of the lateral ventricle are reached at a depth of approximately 0.5 cm in most hemispheres. The approach exposes the hippocampus and the collateral eminence in the floor of the temporal horn. The inferior choroidal point, where the anterior choroidal artery passes through the choroidal fissure, is located near the middle of the exposure. The distance between the inferior limiting sulcus and the inferior choroidal point, just behind the head of the hippocampus, is approximately 11 mm.
Lesions in and around the ambient and proximal quadrigeminal cisterns may prove challenging due to their intimate relationships with important vascular and neuronal structures, including the P2 segment of the PCA, basal vein, and midbrain. The transinsular route directed through the temporal portion of the choroidal fissure can be used for lesions in the posterior crural, ambient, and proximal quadrigeminal cisterns. By opening the choroidal fissure on the forniceal side and elevating the choroid plexus toward the thalamus, the posterior cerebral, anterior, and posterior choroidal arteries as well as the basal vein can be exposed with minimal retraction. The transinsular–transchoroidal approach can be expanded for large lesions extending to the perimesencephalic cisterns by resecting the anterior two thirds of the hippocampus and adjacent parahippocampal gyrus.43 The transsylvian-transinsular-transchoroidal approach may be combined with the pretemporal approach for a more extended exposure of the crural, interpeduncular, and prepontine cisterns.

FIG. 13. Photographs obtained during stepwise dissection of left cerebral hemisphere, showing the relationship between the insular cortex and deep landmarks (A–F). A: Superolateral view of the insula and lateral ventricle. The upper portion of the cerebral hemisphere was removed using an axial cut approximately 1 cm above the posterior ramus of the sylvian fissure. The frontoparietal operculum has been removed to expose the insula lateral to the frontal horn and the body of the lateral ventricle. The anterior portion of the superior limiting sulcus is separated from the frontal horn by the anterior isthmus, and the posterior portion of the limiting sulcus is separated from the atrium by the posterior isthmus. The choroid plexus is attached along the choroidal fissure, located between the fornix and thalamus. The foramen of Monro is located deep to the posterior short gyrus. B: Enlarged lateral view. The M2 branches course along the insula. Most of the branching occurs near the insular pole. The central insular sulcus is the site most replete with insular perforating arteries. C: Posterolateral view of the same hemisphere. The lateral wall of the atrium and the roof of the temporal horn have been removed by dividing the white matter along the inferior limiting sulcus. The posterosuperior insular angle at the junction of the superior and inferior limiting sulci is superficial to the junction of the tail of the hippocampus, calcar avis, and anterior wall of the atrium. The choroid plexus has been elevated to expose the floor of the temporal horn, which is formed by the collateral eminence overlying the collateral sulcus laterally and the hippocampus medially. D: The incision along the inferior limiting sulcus has been extended forward to where the incision begins for the transinsular approach into the temporal horn for a selective amygdalohippocampectomy. The choroidal fissure in the temporal horn has been opened by separating the fimbria of the fornix and choroid plexus to expose the basal vein and PCA in the posterior portion of the ambient cistern below the thalamus. The fimbria of the fornix arises on the surface of the hippocampal formation. The parahippocampal gyrus extends medially below the choroidal fissure. E: The opening along the choroidal fissure has been extended anteriorly without disturbing the attachment of the choroid plexus to the thalamus. The PCA and the basal vein course through the ambient cistern on the medial side of the choroidal fissure. F: The exposure along the choroidal fissure has been extended forward to the back edge of the amygdala. The head of the hippocampus blends into the posterior segment of the uncus. The lateral geniculate body protrudes from the lower surface of the thalamus, just medial to the choroidal fissure and body of the hippocampus, into the upper portion of the ambient cistern. Cal. = calcar; Plex. = plexus; Seg. = segment. See legends to Figs. 1, 2, 4, 5, and 11 for definitions of additional abbreviations.

FIG. 14. Photographs obtained during a cadaveric dissection involving the left transylvian transchoroidal approach to the basal cisterns (A–E). A: Left frontotemporal exposure. The inset shows the position of the head and skin incision for the frontotemporal craniotomy. The sylvian fissure has been opened and the temporal operculum has been retracted to expose the anterior half of the insula. The M2 branches have been retracted to show the position of the incision (dashed black line) along the inferior limiting sulcus. The cortical incision through the inferior limiting sulcus begins approximately 1 to 2 cm posterior to where the anterior end of the long gyrus fuses with the temporal pole and extends posteriorly approximately 1.5 to 2 cm. The anterior one half of the inferior limiting sulcus is devoid of larger perforating arteries. The incision is made lateral to the inferior MCA trunk between the M2 branches, which run parallel to the inferior limiting sulcus. The cortical incision along the inferior limiting sulcus can be located medially, as shown in this photograph, or lateral to the posterior insular vein without sacrificing the vessel. B: The cortical incision, at a depth of approximately 0.5 cm in most hemispheres, provides access to the anterior portion of the temporal horn. The amygdala is located at the anterior edge of the cortical incision in the anterior wall of the temporal horn. This approach exposes the head and body of the hippocampus. The inferior choroidal point, the site at which the anterior choroidal artery passes through the choroidal fissure, is located near the middle of the exposure. The distance between the inferior limiting sulcus and inferior choroidal point is approximately 11 mm. C: The choroidal fissure has been opened by dividing the attachment of the choroid plexus to the fimbria of the fornix, and the choroid plexus has been elevated toward the thalamus. This approach exposes the hippocampus and collateral eminence in the floor of the temporal horn. The upper portion of the medial edge of the parahippocampal gyrus is exposed medial to the hippocampus. Opening the choroidal fissure exposes the branches of the PCA in the ambient cistern. D: The choroid plexus has been retracted to the thalamic side to expose the lateral geniculate body, basal vein, and P2 segment of the PCA. E: The PCA has been elevated to expose the branches of the PCA in the depths of the ambient cistern. Cist. = cistern; Fis. = fissure. See legends to Figs. 1, 2, 4 to 6, 11, and 12 for definitions of additional abbreviations.
Conclusions
The transsylvian route, which requires exposure of part or all of the insula, is commonly selected for lesions in the anterior portion of the cerebral hemisphere, basal cisterns, and skull base. Knowledge of the relationship between the opercula and rami of the sylvian fissure, and the underlying insula can provide a roadmap for approaching insular disease. The partes orbitalis and opercularis and the anterior ascending and horizontal rami can provide reliable landmarks for localizing the sulci and gyri of the insula during sylvian dissection. Opening the frontoorbital ramus, when present, provides direct access to the anterior limiting sulcus and the anteroinferior portion of the insula. The limen recess, the area between the point of entrance of the most lateral LSA and the medial border of the limen insulae, is approximately 1.5 cm wide and is devoid of perforating LSAs. The limen recess may be used as the medial limit of dissection during insular tumor surgery.
A significant contribution of the blood supply to the insula comes from the individual cortical branches and stem arteries of the M2 segment. The largest insular perforating arteries most commonly arise from the central and angular arteries and most often supply the posterior half of the central insular sulcus and the posterior long gyrus.
The venous blood supply of the insula drains predominantly to the deep MCV; however, all of the insular veins may have connections to the SSV. The precentral insular vein, which usually drains the middle short gyrus and insular apex, drains into the SSV more frequently than any other insular vein. Three-dimensional knowledge of neural, arterial, and venous relationships of the insula will assist neurosurgeons in dealing with disorders in this area.
Acknowledgments
We thank Robin Barry and Dustin Harris for assistance in the preparation of figures; David Peace for providing artwork; Ronald Smith, M.S., and Alvero Campero, M.D., of the Microneuroanatomy Laboratory, Department of Neurological Surgery, University of Florida, for their support; and Laura Dickinson for helping with the manuscript.
Contributors: Necmettin Tanriover, MD, Albert L. Rhoton, Jr, MD, Masatou Kawashima, MD, PhD, Arthur J. Ulm, MD, and Alexandre Yasuda, MD
Content from Tanriover N, Rhoton AL, Jr, Kawashima M, Ulm AJ, Yasuda A. Microsurgical anatomy of the insula and the sylvian fissure. J Neurosurg 2004;100:891–922. doi.org/10.3171/jns.2004.100.5.0891.
The Neurosurgical Atlas is honored to maintain the legacy of Albert L. Rhoton, Jr, MD.
References
- Ciszek B, Dabrowska M, Andrzejczak A, et al: Middle superficial cerebral vein. Folia Morphol 57:149–155, 1998
- Cunningham DJ: The development of gyri and sulci on the surface of the island of Reil of the human brain. J Anat Phys 25:338–448, 1890
- Duffau H, Capelle L, Lopes M, et al: The insular lobe: physiopathological and surgical considerations. Neurosurgery 47: 801–811, 2000
- Duffau H, Capelle L, Lopes M, et al: Medically intractable epilepsy from insular low-grade gliomas: improvement after an extended lesionectomy. Acta Neurochir 144:563–573, 2002
- Ebeling U, Kothbauer K: Circumscribed low grade astrocytomas in the dominant opercular and insular region: a pilot study. Acta Neurochir 132:66–74, 1995
- Gibo H, Carver CC, Rhoton AL Jr, et al: Microsurgical anatomy of the middle cerebral artery. J Neurosurg 54:151–169, 1981
- Huang YP, Wolf BS: The basal cerebral vein and its tributaries, in Newton TH, Potts DG (eds): Radiology of the Skull and Brain, Vol 2. St Louis: CV Mosby, 1974, pp 2111–2154
- Krayenbuhl HA, Yasargil MG: Cerebral Angiography, ed 2. London: Butterworths, 1968
- Lang FF, Olansen NE, DeMonte F, et al: Surgical resection of intrinsic insular tumors: complication avoidance. J Neurosurg 95: 638–650, 2001
- Marrone ACH, Severino AG: Insular course of the branches of the middle cerebral artery. Folia Morphol 36:331–336, 1988
- Michotey P, Moscow NP, Salamon G, et al: Anatomy of the cortical branches of the middle cerebral artery, in Newton TH, Potts DG (eds): Radiology of the Skull and Brain, Vol II. St Louis: CV Mosby, 1974, pp 1471–1479
- Naidich TP, Valavanis AG, Kubik S: Anatomic relationships along the low-middle convexity: Part I—Normal specimens and magnetic resonance imaging. Neurosurgery 36:517–532, 1995
- Oka K, Rhoton AL Jr, Barry M, et al: Microsurgical anatomy of the superficial veins of the cerebrum. Neurosurgery 17:711–748, 1985
- Ono M, Kubik S, Abernathey CD: Atlas of Cerebral Sulci. Stuttgart: Thieme, 1990
- Ono M, Rhoton AL Jr, Peace D, et al: Microsurgical anatomy of the deep venous system of the brain. Neurosurgery 15:621–657, 1984
- Özyurt E, Kaya AH, Tanriverdi T, et al: New classification for insular tumors and surgical results of 40 patients. Neurosurg Q 13: 138–148, 2003
- Padget DH: The development of the cranial venous system in man, from the viewpoint of comparative anatomy. Contrib Embryol 36:81–139, 1957
- Penfield W, Faulk ME Jr: The insula: further observations on its function. Brain 78:445–470, 1955
- Rhoton AL Jr: The cerebral veins. Neurosurgery 51 [Suppl 4]: S159–S207, 2002
- Rhoton AL Jr: The cerebrum. Neurosurgery 51 [Suppl 4]: S1–S51, 2002
- Rhoton AL Jr: The lateral and third ventricles. Neurosurgery 51 [Suppl 4]:S207–S272, 2002
- Rhoton AL Jr: The supratentorial arteries. Neurosurgery 51 [Suppl 4]:S53–S120, 2002
- Roper SN, Levesque MF, Sutherling WW, et al: Surgical treatment of partial epilepsy arising from the insular cortex. Report of two cases. J Neurosurg 79:266–269, 1993
- Serrats AA, Vlahovitch B, Parker SA: The arteriographic pattern of the insula: its normal appearence and variations in cases of tumour of the cerebral hemispheres. J Neurol Neurosurg Psychiatry 31:379–390, 1968
- Stein RL, Rosenbaum AE: Normal deep cerebral venous system, in Newton TH, Potts DG (eds): Radiology of the Skull and Brain, Vol 4. St Louis: CV Mosby, 1974, pp 1904–1998
- Suzuki Y, Matsumoto K: Variations of the superficial middle cerebral vein: classification using three-dimensional CT angiography. AJNR 21:932–938, 2000
- Tanriover N, Kawashima M, Rhoton AL Jr, et al: Microsurgical anatomy of the early branches of the middle cerebral artery: morphometric analysis and classification with angiographic correlation. J Neurosurg 98:1277–1290, 2003
- Türe U, Yasargil DCH, Al-Mefty O, et al: Topographic anatomy of the insular region. J Neurosurg 90:720–733, 1999
- Türe U, Yasargil MG, Al-Mefty O, et al: Arteries of the insula. J Neurosurg 92:676–687, 2000
- Umansky F, Juarez SM, Dujovny M, et al: Microsurgical anatomy of the proximal segments of the middle cerebral artery. J Neurosurg 61:458–467, 1984
- Vanaclocha V, Saiz-Sapena N, Garcia-Casasola C: Surgical treatment of insular gliomas. Acta Neurochir 139:1126–1135, 1997
- Varnavas GG, Grand W: The insular cortex: morphological and vascular anatomic characteristics. Neurosurgery 44:127–138, 1999
- Warwick R, Williams PL: Gray’s Anatomy, ed 35. Philadelphia: WB Saunders, 1973
- Wen HT, Rhoton AL Jr, de Oliveira E, et al: Microsurgical anatomy of the temporal lobe: part 1: mesial temporal lobe anatomy and its vascular relationships as applied to amygdalohippocampectomy. Neurosurgery 45:549–592, 1999
- Williams PL: Gray’s Anatomy, ed 38. London: Churchill Livingstone, 1995
- Wolf BS, Huang YP: The insula and deep middle cerebral venous drainage system: normal anatomy and angiography. AJR 90: 472–489, 1963
- Wolf BS, Huang YP, Newman CM: The lateral anastomotic mesencephalic vein and other variations in drainage of the basal cerebral vein. AJR 89:411–422, 1963
- Wolf BS, Huang YP, Newman CM: The superficial sylvian venous drainage system. AJR 89:398–410, 1963
- Yasargil MG: Microneurosurgery. Stuttgart: Georg Thieme Verlag, Vol 1, 1984
- Yasargil MG: Microneurosurgery. Stuttgart: Georg Thieme Verlag, Vol 2, 1984
- Yasargil MG: Microneurosurgery. Stuttgart: Georg Thieme Verlag, Vol 4B, 1996
- Yasargil MG, Çaglar S: Comment, in Varnavas GG, Grand W: The insular cortex: morphological and vascular anatomic characteristics. Neurosurgery 44:127–138, 1999
- Yasargil MG, von Ammon K, Cavazos E, et al: Tumours of the limbic and paralimbic systems. Acta Neurochir 118:40–52, 1992
- Yasargil MG, Wieser HG, Valavanis A, et al: Surgery and results of selective amygdala-hippocampectomy in one hundred patients with non-lesional limbic epilepsy. Neurosurg Clin N Am 4:243–261, 1993
- Zentner J, Meyer B, Stangl A, et al: Intrinsic tumors of the insula: a prospective study of 30 patients. J Neurosurg 85:263–271, 1996
- Zimmerman G, Lewis AI, Tew JM Jr: Pure sylvian fissure arteriovenous malformations. J Neurosurg 92:39–44, 2000
Please login to post a comment.




